metabolismo
- 1. Biokim./Biol.
Organismo bizidunetan gertatzen den erreakzio kimikoen multzoa, substantzien sintesi (anabolismoa) eta degradazioan (katabolismoa) datzana.
- en metabolism
- es metabolismo
- fr métabolisme
1. Biokim./Biol.
-
Berezko erreakzioen energetika -
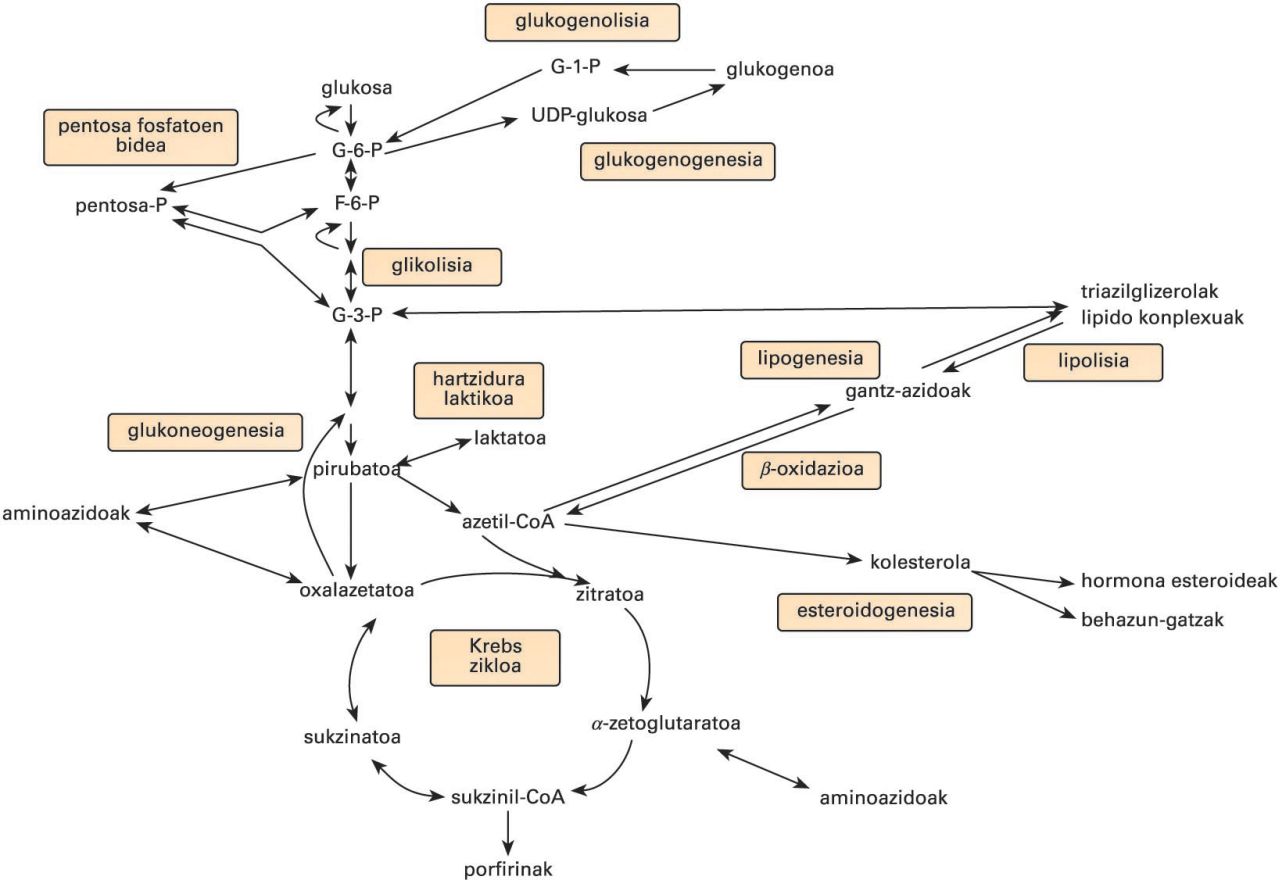
Bide metaboliko nagusiak. G-6-P: glukosa-6-fosfatoa; G-1-P: glukosa-1-fosfatoa; F-6-P: fruktosa-6-fosfatoa; G-3-P: glizeraldehido-3-fosfatoa
- 1. Biokim./Biol.
- Organismo bizidunetan gertatzen den erreakzio kimikoen multzoa, substantzien sintesi (anabolismoa) eta degradazioan (katabolismoa) datzana.
Metabolismoa Edit
Egilea: Olatz Fresnedo
Sarrera eta kontzeptu orokorrak
Metabolismoa bizidunetan gertatzen diren materia- eta energia-eraldaketen multzoa da. Bizidunen propietateak denboran aldatzen ez diren arren, haien funtzioak mantentzea mekanismo molekular konplexu askoren oreka dinamikoan oinarritzen da. Mekanismo horiei esker, bizidunen egitura eta integritatea ingurunearekiko bereiz, mugatuta mantentzen dira, eta aldi berean ingurunearekin materia- eta energia-trukeak bideratzen dira.
Eraldaketen multzoa bide metabolikoetan banatzen da. Konposatu aitzindari baten bidez ekoizkin bat sortzeko beharrezko erreakzio-seriea (hurrenkera) da bide metabolikoa. Adibidez, glukosaren eraldaketaz pirubatoa sorrarazten duen erreakzioen seriea (bidea) glukolisia da. Jatorrizko konposatuak jasaten duen eraldaketa (erreakzio) bakoitza urrats metaboliko bat da, eta kasu gehienetan entzima batek katalizatzen du. Serie horretan sortzen eta eraldatzen diren bitarteko konposatuei bitartekari metaboliko deritze.
Bide metabolikoak bi talde nagusitan sailkatu daitezke: katabolismoa eta anabolismoa.
Erregaia energia zelular bilakatzen duten erreakzioak katabolikoak dira eta katabolismoan biltzen dira.
KATABOLISMOA
erregaiak (karbohidratoak, gantzak) → CO2 + H2O + energia erabilgarria
Energia behar duten erreakzioak —hala nola glukosaren, gantzen edo DNAren sintesikoak— erreakzio anabolikoak dira, eta anabolismoan biltzen dira. Katabolismoan askatutako energiaren forma erabilgarriak konposatu bakunen ondorioz egitura konplexuak ekoizteko erabiltzen dira anabolismoan.
ANABOLISMOA
energia erabilgarria + molekula txikiak → molekula konplexuak
Egoera zelularraren arabera, bide batzuek kataboliko edo anaboliko gisa joka dezakete; era horretako bideak bide anfibolikoak dira.
Animalia-, landare-, onddo- zein bakterio-zelulak eraikitzeko, banakotasunari eusteko eta belaunaldi berriak garatzeko beharrezko mekanismoak funtsean berdin-berdinak dira maila molekularrean. Aldiz, funtsezko osagai molekularrak eraikitzeko materia eta energia-iturri desberdinak erabiltzen dituzte. Erabilitako iturrien arabera, bi talde nagusitan sailkatzen dira: autotrofoak eta heterotrofoak. Inguruneko energia-iturriak desberdinak izanda ere, iturri horien itxuraldaketak (transdukzioak) ezinbestekoak dira biologikoki erabilgarriak diren energia-aldaerak ekoizteko: ATP eta antzeko nukleotido trifosfatoen energia altuko loturak eta koentzima erreduzituen (NAD(P)H eta FADH2) ahalmen erreduzitzailea.
Landareak, adibidez, autotrofoen taldean bildutako organismoen artean daude. Molekula konplexuak eraikitzeko, lurrazaleko konposatu bakun egonkorrak —hala nola ura eta karbono dioxidoa— eta nitrogeno-iturri ez-organikoak erabiltzen dituzte. Horretarako, eguzkitiko argi-energiaz baliatzen dira; fotosintesiko erreakzio fotokimikoen ondorioz energia hori ATP eta NADPH modura gordetzen da, eta, aldi berean, oxigeno molekularra (O2) askatzen da.
Heterotrofoek —gizakiak, adibidez— erabiltzen duten energiaren eta materiaren azken iturriak autotrofoek sintetizatutako biomolekula organikoak dira. Biomolekula horiek bide metabolikoetan erabiltzen dira CO2, H2O eta eratorri nitrogenodun bilakatu arte, eta, aldi berean, energia (ATP eta NAD(P)H) askatzen da. Energia hori zenbait helbururekin erabiltzen da, besteak beste, molekula konplexuak ekoizteko, aitzindari modura ingurunetik hartutako konposatu eta biomolekula bakunak erabiliz.
Erreakzioen energetika eta entzimak
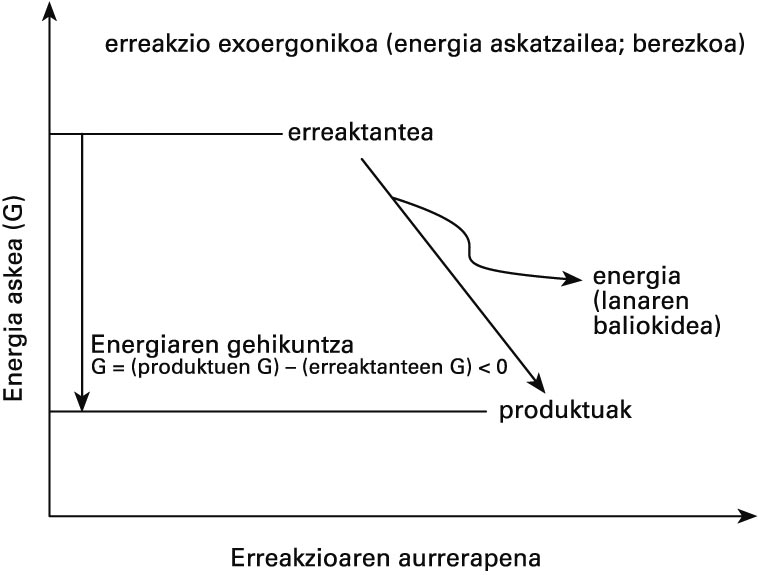
Metabolismoan gertatzen diren eraldaketak bizidunetatik kanpo ere gerta daitezke, eta sistema biologikoetatik kanpo ezinezkoak direnak ezinezkoak dira bizidunen baitan ere: erreakzio bat berez gerta badaiteke, edozein sistema-motatan gertatu ahal izango du. Izan ere, kimika biologikoaren eta ez-biologikoaren funtsezko legeak berberak dira; erreakzioen bideragarritasunaren deskribapen kuantitatiboa egiten duena termodinamika da. Termodinamikaren legeen arabera, erreakzioetan parte hartzen duten konposatuen (erreakzionatzaileen) energia-edukiak zehaztuko du erreakzioa gertatu ahal izango den ala ez. Energia horren adierazle gisa Gibbsen energia askea erabiltzen da. Berez gertatzen diren prozesuetan, Gibbsen energia askearen gehikuntza (aldaketa) negatiboa da, eta haren balioa prozesu horri esker egin daitekeen lanaren neurria da. Bestela esanda, prozesu fisiko-kimiko bat exoergonikoa (energia-askatzailea) baldin bada soilik izango da posible.
Bizidunetan, askatutako energia hori beste prozesu batzuk bultzatzeko erabiltzen da, hala nola berez ezinezkoak diren erreakzioak (endoergonikoak ; energia-erabiltzaileak) bideratzeko, lan mekanikoa egiteko (uzkurdura, higidura zelularra) edo beroa ekoizteko (termogenesia).
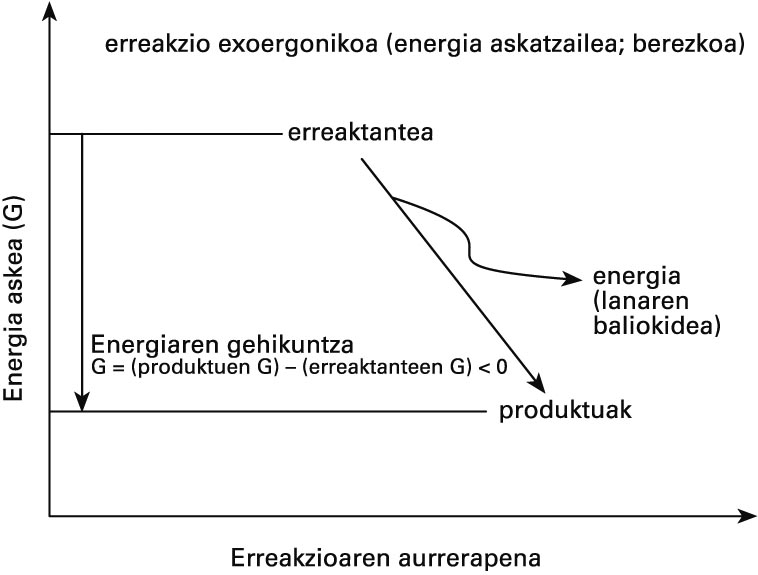
Berezko erreakzioen energetika
Kimika biologikoa eta ez-biologikoa funtsean berdinak diren arren, kimika ez-biologikoan oso geldoa da erreakzio askoren abiadura, eta inguruneko tenperatura, presioa edo pH-a muturrekoak direnean baino ez dira gertatzen. Entzimek oztopo hauek saihesten dituzte: katalizatzaile gisa inguruneko kondizio zorrotzak mimetizatzen dituzten mikroingurune kimiko eta konformazionalak (gune aktiboak) dituzte erreakzioak bideratzeko, eta ondorioz, batetik, sistema biologikoen barne-ingurune leunean erreakzio asko gertatu ahal izateko aukera ematen dute, eta bestetik, zelulen bideragarritasunak ezinbestekoa duen erreakzio-abiadurak lortzen dituzte. Entzimek katalizatzen dituzte metabolismoko eraldaketa kimiko gehienak.
Metabolismoaren erregulazioa
Zelulek, organoek eta organismo osoek mantenurako eta garapenerako behar dituzten erreakzio eta prozesuen multzoak aldakorrak dira. Gainera, inguruneko baliabideak ere, besteak beste, aldatzen diren neurrian, jarduera metabolikoa egokitu egin behar da. Doiketa horiek hertsiki kontrolatzen dira zeluletan.
Entzima erregulatzaileak
Bide metabolikoetan zeharreko fluxuak kontrolatzeko ez da beharrezkoa izaten urrats bakoitzaren erreakzio-abiadura erregulatzea; nahikoa izaten da bide metabolikoaren lehenengo erreakzioren baten abiadura erregulatzea. Urrats hori fluxuaren urrats mugatzailea izango da, eta erreakzioa katalizatzen duena entzima erregulatzaile gisa ezagutzen da. Entzima horien jarduera zenbait modutan erregula daiteke: mekanismo alosterikoen bidez, aldaketa kobalenteen bidez eta beren adierazpen genikoaren aldaketen bidez. Mekanismo horiek espezifikoak dira entzima bakoitzarentzat, eta entzima beraren zereginarekin zerikusia duten faktore eragileek eragindakoak izaten dira. Eragileak zelula barrukoak eta kanpokoak izan daitezke, beharrezko egokitzapenaren arabera.
Konpartimentazioa
Zelula eukariotikoek, mintzez inguratutako organulu espezializatuak dituztenez, prokariotoek baino aukera gehiago dituzte metabolismoa kontrolatzeko. Bide metaboliko bakoitzak zelulan kokagune berezi bat duenez, ingurune eta baliabide espezifikoak izango ditu; horiek, halaber, mintzean zeharreko garraio-sistemen araberakoak izango dira. Konpartimentazioaren abantaila azaltzeko adibide argia da gantz-azidoen metabolismoarena. Zelularen edo organismoaren egoera orokorrak anabolismoa bultzatzen duenean, gantz-azidoak sintetizatzen dira zitosolean, eta bestelako lipido konplexuak sortzen dira (esterifikazio bidez). Aitzitik, egoera katabolikoa bada, hots, energia ekoiztea beharrezkoa bada, gantz-azidoak mitokondrien matrizera garraiatzen dira; han, erregaien eskuragarritasuna handitze hutsagatik oxidatzen dira. Kasu horretan, konposatu baten igorleku metabolikoa haren kokapenaren araberakoa da, eta mintzean zeharreko fluxuaren erregulazioaren mende dago.
Organoen espezializazio metabolikoa
Goi-mailako eukariotoetan ehunen zeregin fisiologikoaren berezitasunek (espezializazio tisularrak) aitzindari metabolikoen eta energiaren beharrizan eta gaitasun metaboliko espezifikoak zehazten dituzte. Ondorioz, ehun bakoitzak diseinu metaboliko eta egokitzapen bereziak ditu. Guztien koordinazioa ezinbestekoa da organismoaren erantzunak integralak izan daitezen. Koordinazio horren arduradun nagusiak nerbio-sistema eta sistema hormonala dira.
Bide metabolikoak
ATP-ekoizpena
Azido trikarboxilikoen zikloa (edo Krebsen zikloa). Azetil-A koentzimaren (azetil-CoA) azetiloaren oxidazio osoa (CO2 arte) gertatzen da bide honetan. Azetil-CoAren iturri nagusiak hiru dira: karbohidratoen glukolisiz askatutako pirubatoa, gantz-azidoak eta aminoazido zetogenikoak. Bai azetil-CoA lortzea bai azido trikarboxilikoen zikloa mitokondrian gertatzen dira, eta prozesu horietan gertatzen diren oxidazio-erreakzioetan FAD eta NAD+ dira elektroi-hartzaileak (erreduzitzen diren koentzimak). Hartzaile horien aldaera erreduzituak (FADH2 eta NADH) fosforilazio oxidatzailean erabiltzen dira ATPa ekoizteko. Azido trikarboxilikoen zikloko bitartekariak, gainera, zenbait biosintesi-prozesutan erabiltzen dira; horregatik, bidea anfibolikoa dela esan daiteke.
Fosforilazio oxidatzailea. ATPa ekoizteko mitokondriako arnas kateko elektroi garraioaz baliatzen den mekanismoa da. Elektroi horien jatorria karbohidratoen eta gantz-azidoen gisako erregaiak dira; elektroiak arnas katera transferitzen dituzten konposatuak NADHa eta FADH2 a dira, eta azken hatzailea oxigeno molekularra (O2) da. Barne-mintz mitokondrialean kokatutako proteina-konplexu integralen multzoa da arnas katea. Haren jarduerak hidrogeno-ioiak (H+) higiarazten dituen indarra sorrarazten du (gradiente elektrokimikoa mitokondriaren bi mintzen arteko espazioaren eta matrizaren artean). Indar horrek bultzatzen du ATP sintasaren katalisia (ADParen fosforilazioa) ATPa ekoizteko.
Fotosintesiaren fase argiko erreakzioak. Goi-mailako landareetan, auketan eta zianobakterioetan, fase argiko erreakzioetan argia xurgatzeari esker, ur molekuletatik elektroiak askatzen dira (oxidatu), eta zenbait elektroi-garraiatzailetan garraiatuta, NADP+ a erreduzitzeko erabiltzen dira. Elektroiekin batera, eta haien garraioari esker, ur molekuletatik askatutako hidrogeno-ioiek gradiente elektrokimiko bat sorrarazten dute; gradiente hori ATPa ekoizteko erabiltzen da (antzera gertatzen da fosforilazio oxidatzailean ere). Ondorioz, O2-a askatzen da.
Karbohidratoak
Glukolisia. Bide honetan, glukosa pirubatoa ekoizteko eraldatzen da 10 urratsetan. Glukosaz gain, fruktosa, galaktosa eta antzeko beste monosakarido batzuk ere erabil daitezke glukolisian. Prozesuaren helburuak hexosa horien katabolismoa hastea, aitzindari biosintetikoak ekoiztea eta energia (ATP eta NADH) askatzea da. Metabolismo anaerobikoan (hartzidura laktikoa eta alkoholikoa) energia lortzeko bide nagusia da. Prozesuaren erregulazioak zeregin horiei eta organismo osoaren glukosa-beharrizan / glukosa-eskuragarritasun erlazioari erantzuten dio.
Glukoneogenesia. Bereziki garrantzitsua da baraualdietan, laktatoa, glizerola, alanina eta antzeko aitzindari ez-glizidikoen ondorioz glukosa ekoizten duelako. Hiru erreakziotan (itzulezinak direnak) ez beste guztietan, prozesu hau glukolisiaren kontrakoa da, pirubatotik hasita. Glukoneogenesiaren gune garrantzitsuena gibela da; organo horrek, beste askok ez bezala, zeluletatik glukosa odolera kanporatzeko gaitasuna dauka (glukosa 6-fosfatasa entzima daukalako), eta horregatik, funtsezkoa da glukosaren homeostasiarako (gluzemia-mailari eusteko). Haren erregulazioa glukolisiarenaren kontrakoa da.
Glukogenoaren metabolismoa. Glukogenoa animalien ehun guztietan (bereziki, gibelean eta muskuluan) eta zenbait bakteriotan eta legamiatan glukosa metatzeko erabiltzen den polisakaridoa da. Gizakian, haren sorrera eta erabilera, gibelean bereziki, gluzemiaren (odoleko glukosa-kontzentrazioaren) araberakoak dira.
Calvinen zikloa eta pentosa fosfatoen bidea. Calvinen zikloa goi-mailako landare gehienen kloroplastoetan gertatzen da: fotosintesiaren fase iluneko erreakzioei dagokie; fase argiko erreakzioetan sortutako ATPa eta NADPHa erabilita glukosa ekoizten du, CO2-aren finkapenaren ondorioz. Calvinen zikloan, Rubisco laburduraz ezaguna den entzimak (karbono dioxidoaren finkatzailea da) eta pentosa fosfatoen bideko eta glukolisiko hainbat entzimak hartzen dute parte. Pentosa fosfatoen bidea glukosaren oxidaziorako aukerako bidetzat har daiteke; NADPH (koentzimaren forma erreduzitua) eta pentosa fosfatoak (bereziki, erribosa fosfatoa, nukleotidoen osagaia) ekoiztea da haren funtzioa. Gai horien beharrizanen arabera, bideak diseinu metaboliko desberdinak ditu.
Lipidoak
Lipolisia eta gantz-azidoen oxidazioa. Lipolisi terminoa modu generikoan lipidoen apurketa dela uler daitekeen arren, metatutako azilglizerolen hidrolisi entzimatikoa adierazteko erabiltzen da. Askatutako gantz-azidoak oxidatu egiten dira energia lortzeko. Oxidaziorako hainbat bide daude; gizakian garrantzitsuena mitokondriako beta-oxidazioa da. 4 erreakzioko zikloa behin eta berriro errepikatzen da, eta ziklo bakoitzean kate luzeko gantz-azidoaren 2 karbono azetil-CoA moduan askatzen dira. Aldi berean, NADH eta FADH2 (forma erreduzituak) lortzen dira, eta fosforilazio oxidatzailean erabiltzen dira horiek.
Lipogenesia. Modu generikoan edozein lipidoren eraketa adierazten badu ere, gantz-azidoen biosintesia adierazteko erabiltzen da. Gainera, sintesi hori gertatu ahal izateko sortutako gantz-azidoak erabili egin behar direnez —bereziki, triazilglizerolak eratzeko—, azetil-CoA-tik abiatuta triazilglizerolen sorrerarako prozesuak adierazten ditu lipogenesi terminoak. Gantz-azidoen sintesia zitosolean gertatzen da, errepikatzen diren 4 erreakziori esker. Ziklo bakoitzean, gantz-azidoa 2 karbonorekin luzatzen da, eta NADPHak jokatzen du erredukzio-erreakzioen elektroi-emaile gisa. Erregai-soberakinak daudenean gertatzen da prozesua.
Lipido konplexuen eta esteroideen sorrera. Lipido konplexuak ekoizteko, lipidoen osagaiak elkartu egiten dira, batez ere transferentzia-erreakzioen bitartez. Horretarako, elkartuko den osagaietakoren batek aktibatuta egon behar du, CDP edo A koentzima lotua duela. Esteroideei dagokienez, esterolak dira arruntenak; aitzindari nagusitzat azetil-CoA erabilita sintetizatzen dira.
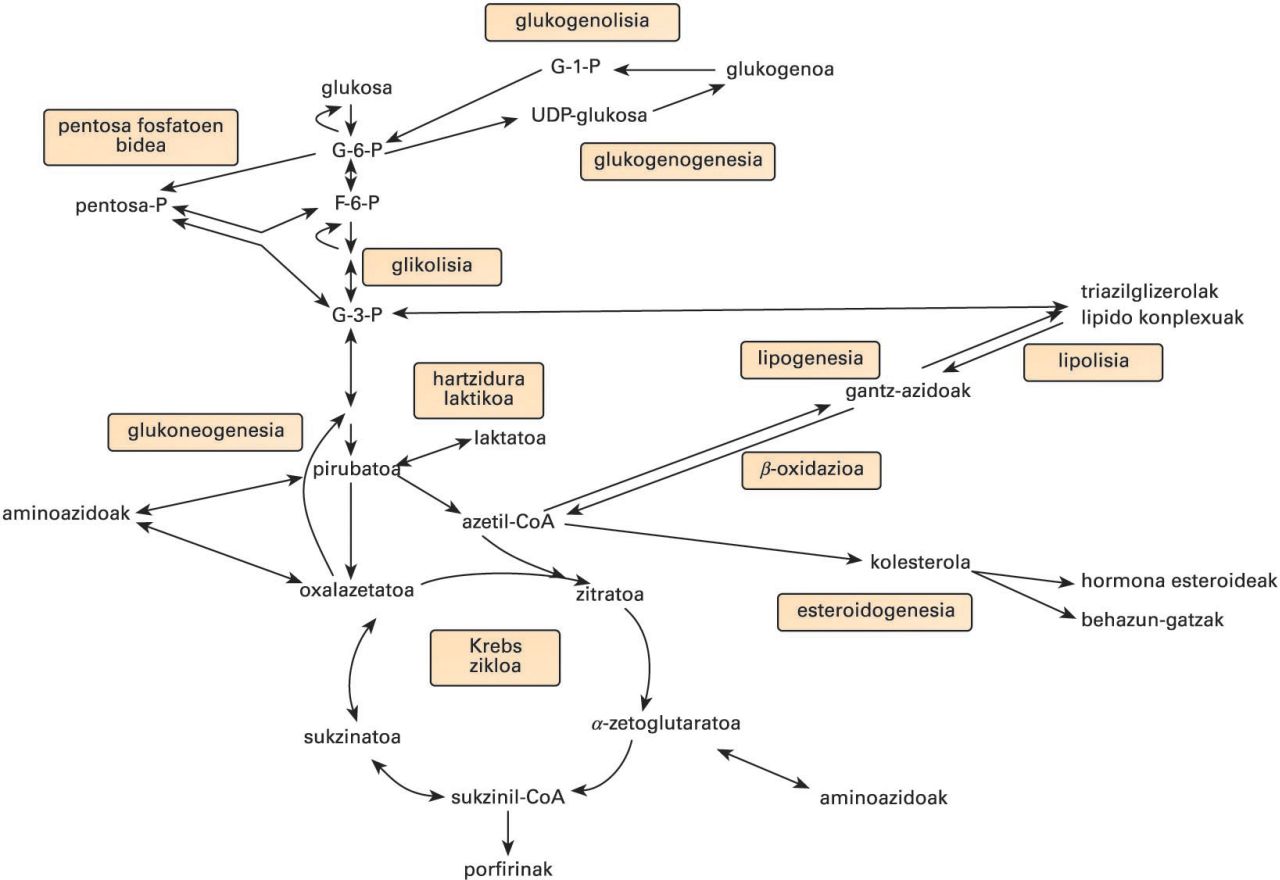
Bide metaboliko nagusiak. G-6-P: glukosa-6-fosfatoa; G-1-P: glukosa-1-fosfatoa; F-6-P: fruktosa-6-fosfatoa; G-3-P: glizeraldehido-3-fosfatoa
Konposatu nitrogenodunak
Proteinen eta aminoazidoen katabolismoa. Zeluletako proteinak berriztatzeko, proteina horiek hidrolizatu egiten dira eta beren osagai diren aminoazidoak askatzen dituzte. Aminoazido horien katabolismoaren lehen urratsa, gehienetan, alfa-karbonoko amino taldea kentzea izaten da; aminoazido desberdinetan antzeko moduan gertatzen da prozesu hori. Amino taldeak iraizteko bideak aldatu egiten dira izaki batzuetatik besteetara; gizakiaren kasuan, urea modura kanporatzen da. Amino taldea askatzen denetik aurrera, aminoazido bakoitzaren bide katabolikoa berezia da; kasu askotan, glukosa edo azetil-CoA ekoizteko erabiltzen dira aminoazido horiek.
Aminoazidoen biosintesia. Aitzindari modura bitartekari arruntak erabiltzen dira; adibidez, Krebsen ziklokoak. Aminoazido bakoitzak sintesirako bide espezifiko bat duen arren, azkeneko erreakzioa berdina izaten da: alfa-aminoa gehitzea. Amino talde hori glutamatoak eta glutaminak ematen dute
Nukleotidoen biosintesia eta degradazioa. Aitzindari ez-nukleotidikoen ondoriozko biosintesia (de novo delakoa), normalean, zelulen bikoizketaren aurretik gertatzen da, DNA sortzeko. Azido nukleikoak (RNA eta DNA) berriztatzen direnean, nukleotidoak ez dira guztiz katabolizatzen. Azido nukleikoen osagai diren base nitrogenatuak (bai purinak, bai pirimidinak) berrerabili egiten dira nukleotidoak sortzeko. Izan ere, beharrezkoa izanez gero katabolizatu egiten dira base nitrogenatuak, eta katabolismo horren abiaduraren neurri berean izaten da beharrezkoa de novo sintesia.