proteina
- 1. Biokim.
Lotura peptidikoz lotutako aminoazidoez osatutako makromolekulen izen generikoa. Bizidun guztien osagai dira.
1. Biokim.
-
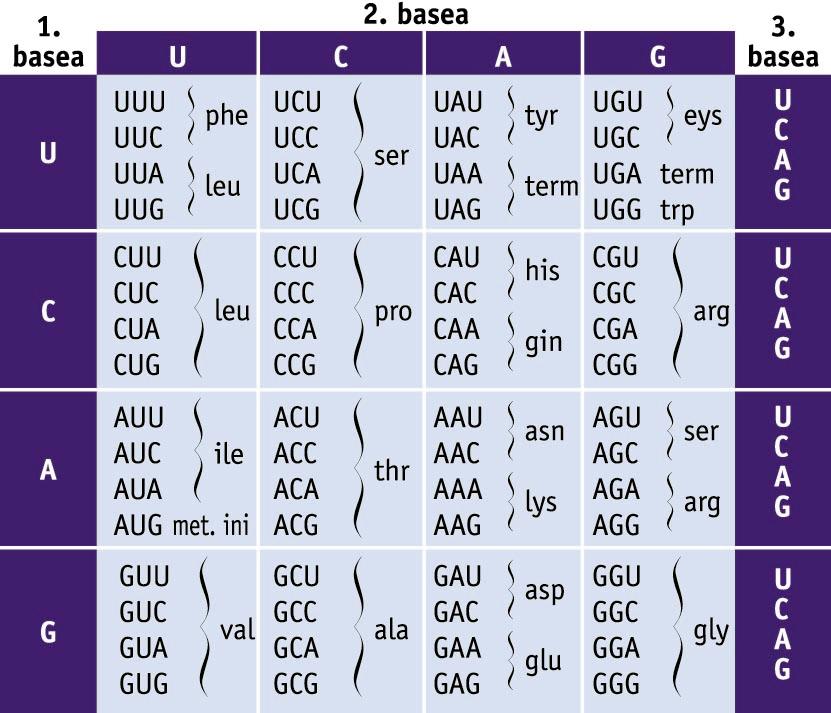
Kode genetikoa. 64 kodoiak eta kodetzen dituzten aminoazidoen taula -

Proteinen zenbait funtzio. a) Keratina ornodun guztiek ekoizten duten proteina da, eta ile, adar, artile, oskol, azazkal eta lumetako osagarri nagusia da. b) Ipurtargiek sortzen ... -
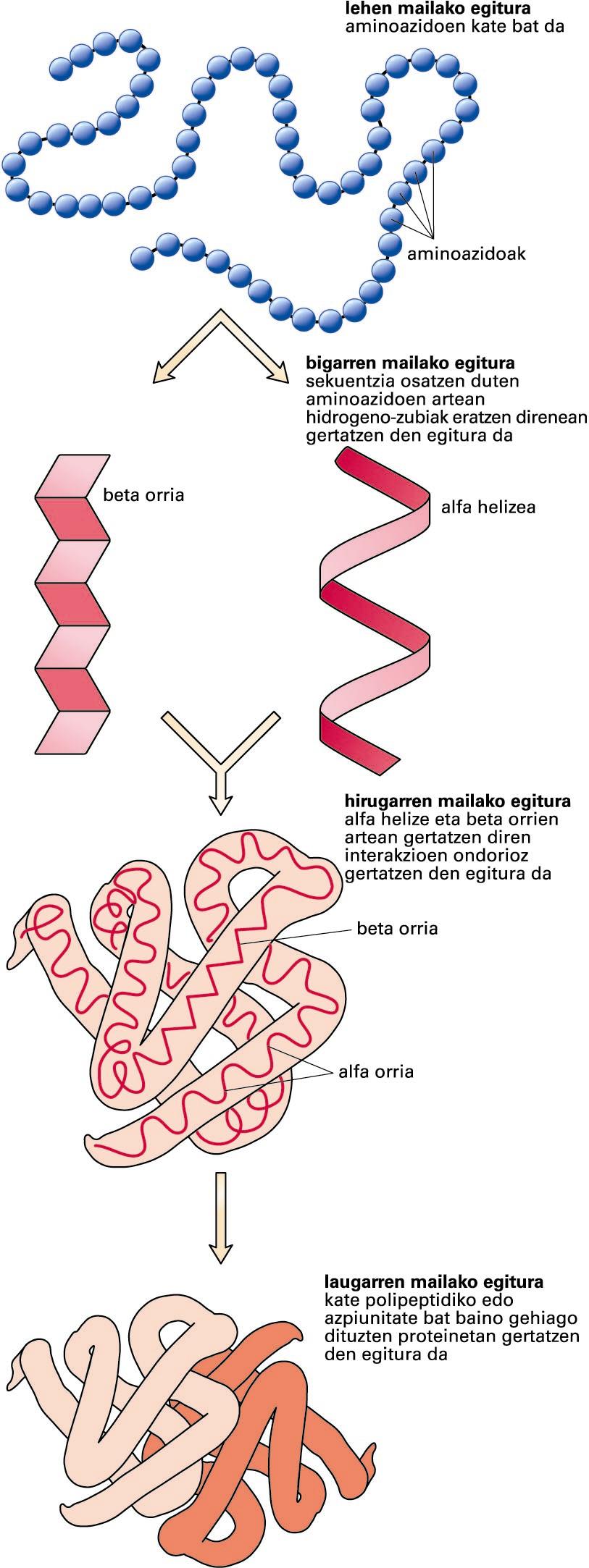
Proteinen egituraren lau mailak -
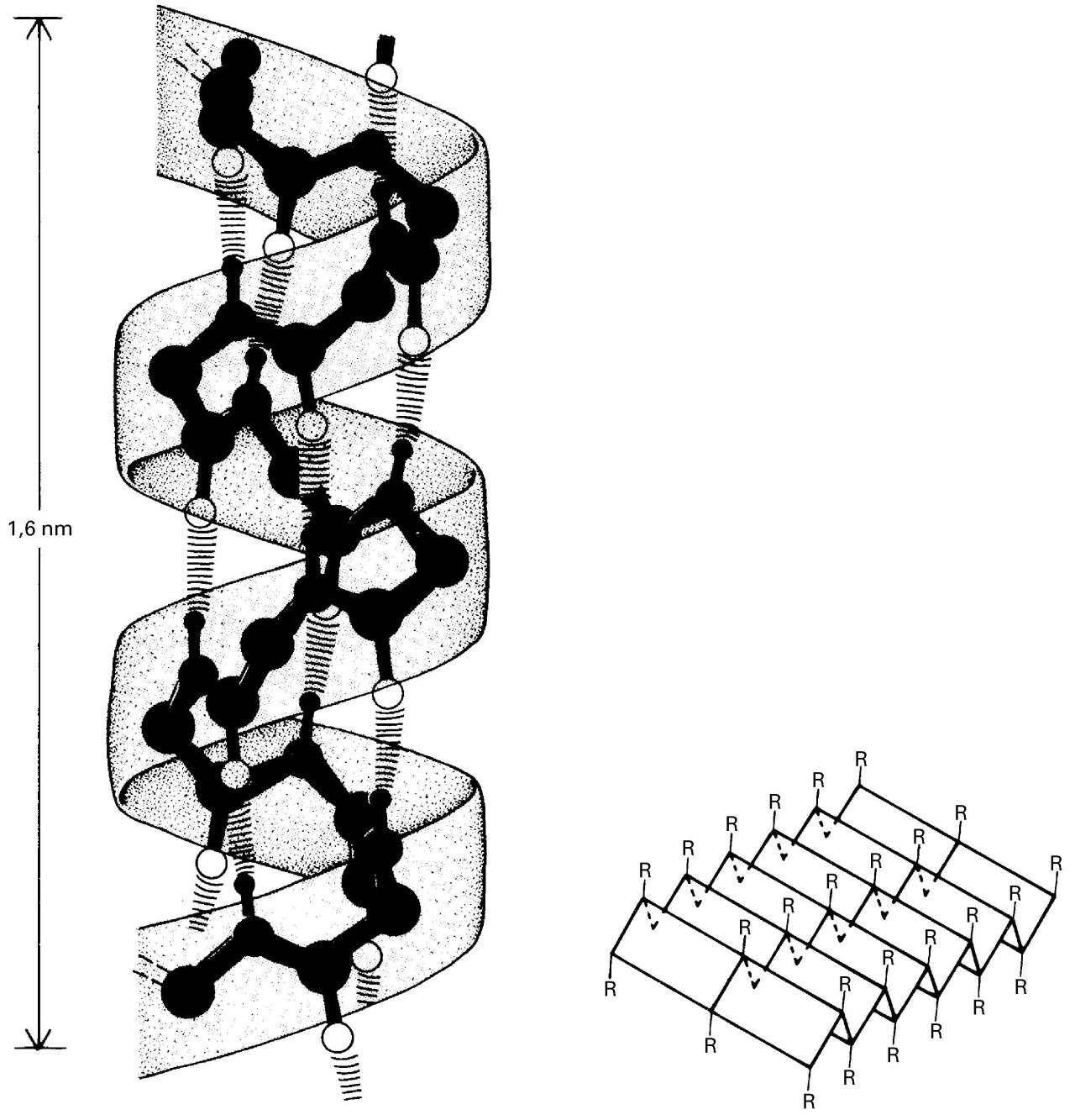
Bigarren mailako egituraren α helizea (ezkerrean) eta β orria (eskuinean) -
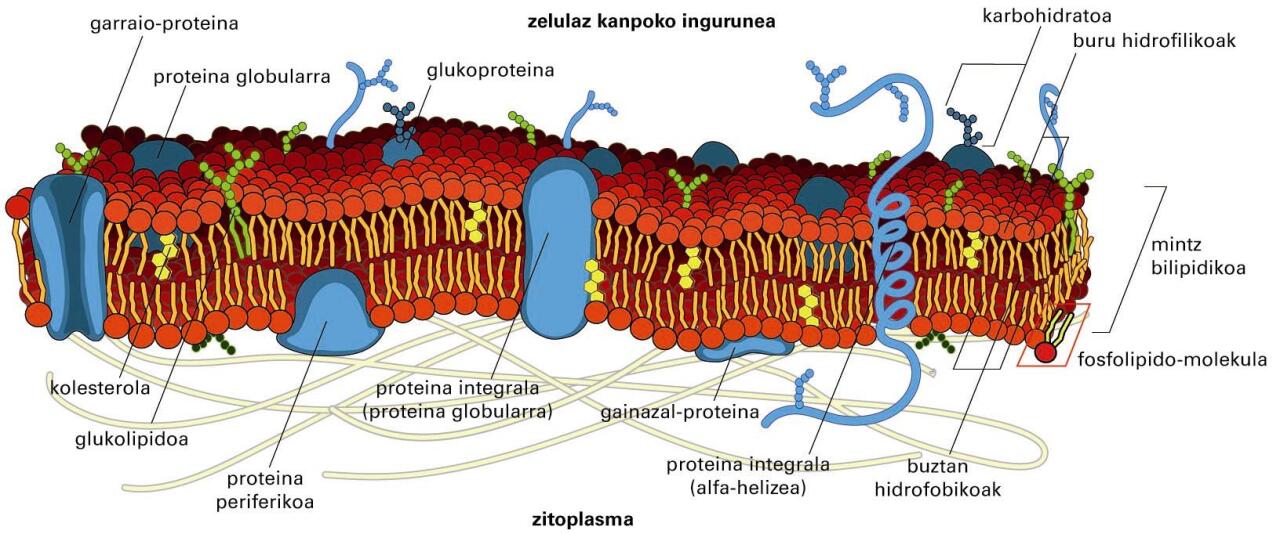
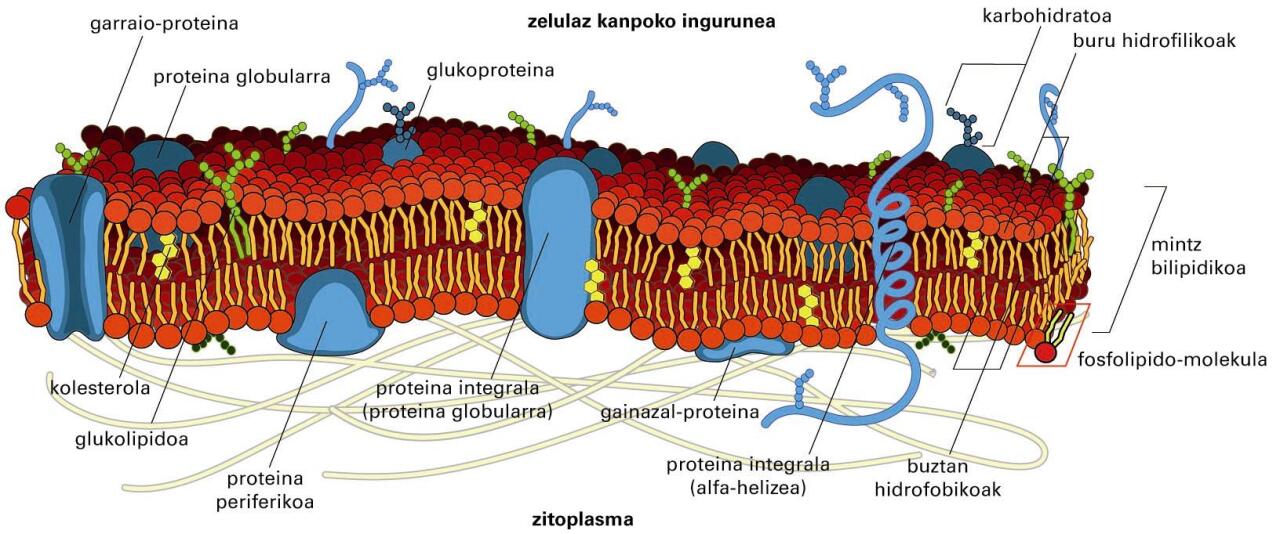
Mintzeko proteinak
- 1. Biokim.
- Lotura peptidikoz lotutako aminoazidoez osatutako makromolekulen izen generikoa. Bizidun guztien osagai dira.
Proteina Edit
Egilea: Miren Josu Omaetxebarria
Proteinak aminoazidoz osatutako konposatu organikoak dira. Aminoazidoak euren karboxilo taldeen eta aminoen artean gertatzen diren lotura peptidikoen bidez lotzen dira, eta kate luzeak osatzen dituzte. Katea proteina litzateke, eta katebegi bakoitza aminoazido bat.
Kode genetikoa. 64 kodoiak eta kodetzen dituzten aminoazidoen taula
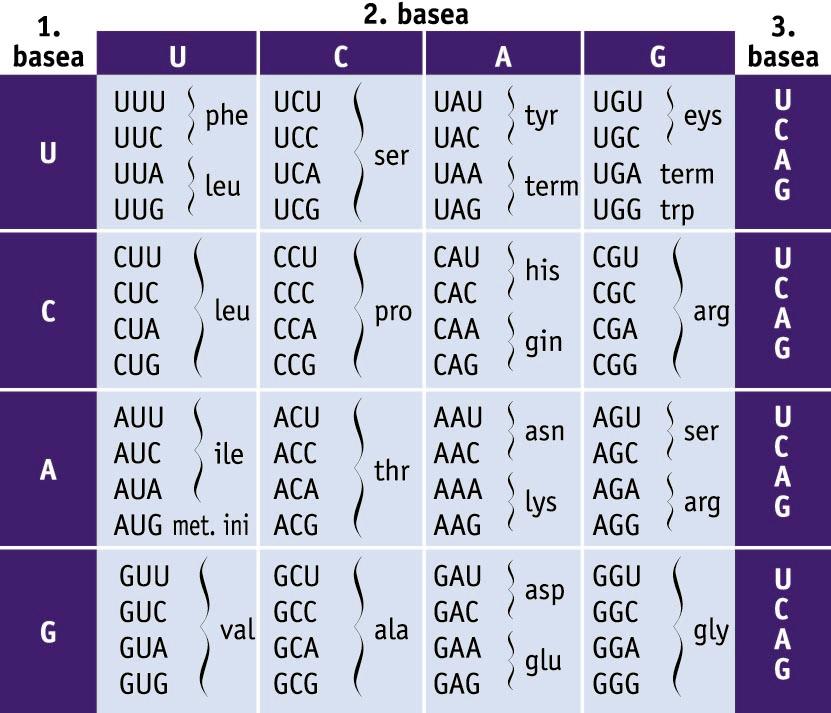
Proteina bakoitzak aminoazido-sekuentzia jakin bat du. Aminoazido-sekuentzia jakin hori geneetan gorderik dagoen informazioak zehaztuko du. Geneetako informazioa kode genetikoan oinarritzen da. Kode genetikoa hiru nukleotido zenbait eratan konbinatuta osatutako hirukoteen bilduma da. Nukleotido-hirukoteari kodoi deritzo, eta kodoi bakoitzak aminoazido bat adieraziko du; esate baterako, AUG metionina aminoazidoaren kodoia da (ikus taula).
Proteina guztiak, bai bakterioenak bai izaki konplexuenenak, hainbat konbinaziotan kobalenteki lotutako 20 aminoazidoz osatutako kateak dira. Oinarrizko 20 aminoazido horiek proteinen sekuentzia idazteko alfabetoa osatzen dutela esan dezakegu. Zeluletan 20 aminoazido berberak konbinazio eta sekuentzia desberdinetan lotuko dira, eta ezaugarri eta funtzio zeharo desberdinak izan ditzaketen proteinak osatuko dituzte. Proteinak, beste makromolekulen gisara —hala nola polisakaridoen eta azido nukleikoen gisara—, izakien oinarrizko osagaiak dira, eta ezinbestekoak dira zelula barneko prozesuetarako. Proteina-mota asko dago, eta zelula bakar batean milaka proteina-mota aurkitu daitezke, peptido txikietatik hasi eta polimero handietara. Proteina asko erreakzio biokimikoak katalizatzen dituzten entzimak dira. Beste batzuek funtzio mekanikoa dute —esaterako, giharretako miosinak— edo egituretan agertzen dira —zitoeskeletoa osatzen duen aktina adibidez, zeina ezinbestekoa baita zelulen forma mantentzeko—. Beste zenbait funtsezkoak izango dira seinalizazio-zidorretan, immunitate-erantzunean, edota ziklo zelularrean. Besteak beste entzimak, hormonak, antigorputzak, muskulu zuntzak, larruazala, lumak, amaraunak, onddo eta sugeen pozoiak eta beste hainbat funtzio eta aktibitate izan ditzaketen makromolekulak dira proteinak.

Proteinen zenbait funtzio. a) Keratina ornodun guztiek ekoizten duten proteina da, eta ile, adar, artile, oskol, azazkal eta lumetako osagarri nagusia da. b) Ipurtargiek sortzen duten argia luziferina proteinak eta ATPak elkarrekin erreakzionatzean gertatzen den eta luziferasa entzimak katalizatzen duen erreakzio baten ondorioa da. c) Globulu gorri edo eritrozitoek oxigeno-garraiatzailea den hemoglobina ugari dute
Proteinak oinarrizkoak dira animalien dietan ere; izan ere, aipatutako oinarrizko 20 aminoazidoetatik batzuk baino ezin dituzte sintetizatu animaliek, eta ondorioz, elikagaietatik lortu behar dituzte gainerako aminoazidoak. Digestioan proteinak apurtzean askatzen diren katebegiak, hau da, aminoazidoak, metabolismoan erabiliko dira.
Proteina hitza prota hitz grekotik dator, eta “funtsezkoa” adierazten du. Jöns Jakob Berzelius kimikari suediarrak deskribatu eta aipatu zituen lehenengoz proteinak 1838an. Baina 1926an James B. Sumnerrek ureasa entzima proteina bat zela erakutsi zuen arte, ez zen aintzat hartu proteinek izakietan zuten eginkizun garrantzitsua. 1953an, Frederick Sangerrek inoiz sekuentziatutako lehen proteinaren —intsulinaren— sekuentzia argitara eman zuen, eta lorpen horri esker kimikako Nobel saria irabazi zuen 1958an.
Proteinen sintesia
Proteinak edo RNA-molekulak sintetizatzeko informazioa gordetzen duen DNA-zatiari gene deritzo. Hala ere, DNAren informazio genetikoa ez da zuzenean erabiliko proteinen sintesirako. RNA mezularia (mRNA) da bitartekari geneetako informazioa proteina funtzionala izango den aminoazido-sekuentzia bihurtzeko. DNA-molekula bat moldetzat hartuta sintetizatuko den molekula osagarria da RNA. DNAren menpeko RNA-sintesiari transkripzio deritzo. RNA polimerasa izeneko entzimak DNA-sekuentzia kopiatuko du, eta hala, RNA nukleotido-kate osagarria sintetizatuko da. DNAren eta RNAren arteko desberdintasunen artean aipatzekoa da RNAren sekuentzian timinen ordez uraziloak daudela. Horretaz gain, RNAren nukleotidoetako azukrea erribosa da, eta DNAn, berriz, azukrea desoxirribosa da (oxigeno atomo bat gutxiago).
Proteina jakin bat sintetizatzeko informazioa duen mRNA eta sintesirako beharrezkoak diren RNA erribosomikoa (rRNA) eta RNA garraiatzailea (tRNA) ekoitziko dira transkripzioan. Proteinen sintesia erribosometan gertatzen da, eta erribosoma horien osagai nagusia da rRNA. mRNA, berriz, mezu genetikoa DNAtik proteinen sintesirako makineriara (erribosometara) garraiatuko duen bitartekaria da. mRNA hori izango da erribosomek proteina sintetizatzeko erabiliko duten molekula-moldea. mRNAren nukleotido-hirukoteek kodoiak osatzen dituzte, eta kodoi horiek bata bestearen ondoren irakurriko dira; kodoi bakoitzari aminoazido bat dagokio. mRNA lau nukleotidoren (adenina, uraziloa, guanina eta zitosina) konbinazioz osatutako polimeroa izanik, nukleotido-hirukote edo kodoi ezberdin posibleen kopurua guztira 43 da, hau da, 64. Beraz, oinarrizko 20 aminoazidoak kodetzeko 64 kodoiz dago osatuta kode genetikoa. Hori horrela izanik, badira kodoi bakar batek kodetutako aminoazidoak, hala nola metionina (AUG) eta triptofanoa (UGG); beste muturrean dago, besteak beste, leuzina aminoazidoa, zeina sei kodoi desberdinek (UUA, UUG, CUU, CUC, CUA eta CUG) kodetzen baitute. Hori hala izanik, kode genetikoa endekatuta dagoela esan ohi da. Kodoiaren lehen bi nukleotidoek zehazten dute askotan aminoazidoa, eta hirugarren nukleotidoa aldakorra da eta kulunkan dagoela esan ohi da.
mRNAk zuzendutako proteinen sintesiari itzulpen deritzo. Itzulpena lau fasetan gertatzen da: aktibazioa, hasiera, luzapena eta amaiera. Sekuentzia jakineko polipeptido baten sintesia gerta dadin, ezinbestekoa da bi baldintza betetzea. Batetik, aminoazido bakoitzaren karboxilo taldeak aktibatuta egon behar du ondoko aminoazidoarekin lotura peptidikoa gerta dadin; eta bestetik, eransten den aminoazido bakoitzak bat etorri behar du mRNAn jasotako informazioarekin. Itzulpena gerta dadin, erabakigarria izango da aminoazidoak tRNAri lotzea. Erreakzio horri aminoazilazio deritzo, eta zitosolean gertatzen da (ez erribosometan). 20 aminoazidoetako bakoitza, ester lotura bidez, espezifikoki lotuko zaio tRNAri; horretarako, aminoazil-tRNA sintetasa entzimak ATParen energia erabiliko du. tRNA jakin baten antikodoiaren arabera, uste izatekoa da tRNA horri aminoazido jakin bat lotuko zaiola; baina kode genetikoan aminoazido bera kodetzen duten kodoi bat baino gehiago dagoenez, antikodoi desberdinak dituzten tRNAek aminoazido bera garraia dezakete. Aminoazidoa lotuta daraman tRNA zamatuta dagoela esaten da, eta aminoazil-tRNA deritzo. Sintetizatuko den polipeptidoaren kodea daraman mRNA erribosomaren azpiunitate txikiari eta aminoazil-tRNA hasleari lotuko zaie. Ondoren, erribosomaren azpiunitate handia batuko da, eta hasierarako konplexua eratuko da. Lehen aminoazil-tRNA (metionina darama) mRNAren AUG kodoiari lotuko zaio —polipeptidoaren hasiera adierazten du kodoi horrek—; prozesu horretan, GTPak eta hasiera-faktoreek hartuko dute parte. Luzapen-fasean, sortu berria den kate polipeptidikoa hazten joango da. Aminoazil-tRNAek erribosomara garraiatuko dute lotua duten aminoazidoa, eta dagokien mRNA kodoiarekin parekatuko dira; hala, aminoazido bat bestearen atzean kokatu eta lotuko da, eta katea luzatu egingo da. Kate polipeptidikoari aminoazidoak erantsi ahala gertatzen den GTP hidrolisiak bideratuko du mRNAren gaineko erribosomaren desplazamendua. Proteinak, beti, amino muturretik hasi eta karboxilo muturrerantz biosintetizatzen dira. Bukaera, erribosomak stop kodoi bat (UAA, UAG edo UGA) aurkitzen duenean gertatuko da. Ez dago kodoi hori ezagutuko duen tRNArik, baina askapen-faktore batzuk ezagutu ditzakete zentzugabeko kodoi horiek, eta kate polipeptidikoa erribosomatik askatzea eragingo dute. Sintetizatutako edozein proteinaren neurria kate polipeptidikoak dituen aminoazido-kopuruaren arabera eman daiteke, edo haren masa molekular osoaren arabera; masa molekular osoa daltonetan (Da) adierazten da. Sintetizatutako polipeptidoek erribosometatik askatu aurretik edo ondoren tolestu eta hirugarren mailako konformazioa hartzean, proteinaren egitura aktiboa osatuko dute.
Aipatu bezala, mRNAren itzulpena zitosoleko erribosometan gertatzen da. Sintetizatutako proteina zitosolekoa ez beste edozein organulutakoa bada, hara garraiatuko da. Proteina bakoitza dagokion zelulako atalera edo zelulatik kanpora garraiatuko da. Proteinak organulu jakin baten barnealdera, organuluaren mintzera, zelularen kanpo-mintzera edo zelulatik kanpora bideratuko dira. Bideratzea era egokian gertatzea ezinbestekoa da zelularentzat, bideratze-akatsek gaixotasunak eragin baititzakete. Proteina bera da dagokion lekura bideratua izan dadin beharrezkoa den informazioaren jabe. Informazioa kate polipeptidikoan dago. Jarraian dauden zenbait aminoazidoz osatutako ilarari peptido-seinale deritzo. Peptido-seinaleak amino-muturrean (zenbaitetan, karboxilo-muturrean) egoten dira, eta 6-136 aminoazido oinarrizko eta hidrofoboz osatutako sekuentziak dira. Peptido-seinalea duten proteinak nukleora, mitokondrietara edo erretikulu endoplasmatikora, kloroplastoetara edo peroxisometara joango dira, besteak beste. Peptido-seinaleaz gain, proteinek jasan ditzaketen itzulpen ondorengo aldaketek, glikosilazioak esaterako, proteinen bideratzea eragin dezakete.
Nahiz eta zelulek milaka proteina izan, sintesia momentuko beharrizanen arabera gertatuko da. Zelula bateko milaka proteinaren sintesia estuki erregulatuta dagoen prozesua da, eta, kondizioen arabera, beharrezkoak diren proteinak eta kopuruak baino ez dira sintetizatuko.
Batzuetan, gerta daiteke proteina akastunak sintetizatzea, edo sintetizatu ondoren hondatzea, estres oxidatiboaren ondorioz adibidez. Zelulak birziklatu egingo ditu proteina akastun horiek. Gainera, proteinek biziraupen desberdinak izan ditzakete; ondorioz, batzuk luzaroan mantendu daitezke, baina beste batzuk suntsitu, eta berriro sintetizatu beharko dira. Eukariotoen kasuan, ubikitina izango da proteinen birziklapenean parte hartuko duen proteina. Haren funtzio nagusietariko bat suntsitu beharreko proteinak proteolisirako markatzea da. Zenbait ubikitina-molekula lotuko zaizkio suntsitu behar den proteinaren lisina bati poliubikitinazio izeneko prozesuan. Proteina ubikitinatu ondoren, markatuta dagoela, proteasomara zuzendu, eta han proteolisia (proteinaren lisia, apurketa) gertatuko da.
Une edota baldintza jakin batzuetan zelula-mota jakin batean oro har dauden proteinen bildumari proteoma deritzo; proteinak eta proteomak ikertzen dituen zientziaren atalari proteomika deritzo.
Proteinen egitura
Biologikoki aktiboa izateko, zelulan sintetizatu berri den kate polipeptidikoa egitura tridimensionala hartuz tolestuko da. Tolestea gertatu aurretik edo ondoren, kate berriak aldaketa entzimatikoak jasan ditzake, hala nola aminoazido bat edo gehiago kentzea (gehienetan amino muturrekoa), aminoazido jakin bat edo batzuei azetilo, fosfato, metilo, karboxilo edo beste talderen bat gehitzea, mozketa proteolitikoa edota oligosakarido edo talde prostetikoren bat lotzea. In vivo gertatzen den tolestea itzulpena gertatu ahala hasten da maiz, hau da, proteinaren amino muturra tolesten hasiko da sintesia bukatu aurretik eta karboxilo muturra oraindik erribosomari lotuta dagoela. Hiru dimentsioko egitura aminoazido-sekuentziaren araberakoa da. Katea osatzen duten aminoazidoek berezko ezaugarri kimikoak dituzte. Aminoazidoak hidrofiloak, hidrofoboak edo elektrikoki kargatuak izan daitezke, besteak beste. Elkarrekin eta zelula barneko ingurunearekin interakzionatuko dute (hidrogeno-zubiek, interakzio ionikoek, Van der Waalsen indarrek edota interakzio hidrofoboek bideratuta), eta ondorioz, kate polipeptidikoak ondo zehaztutako hiru dimentsioko egitura hartuko du. Proteina bat espontaneoki tolestean hartzen duen forma funtzionalari proteinaren egoera natibo deritzo.
Proteina gehientsuenetan, aminoazido polarrek edo kargadunek interakzionatuko dute inguruneko ur-molekulekin; aminoazido hidrofoboek, berriz, muin hidrofobo bat osatzen dute, eta hala, tolestura egonkorragoa da. Hain zuzen ere, badirudi aminoazido hidrofoboak inguruko ur-molekuletatik ezkutatze horretan datzala tolesteko prozesua.
Nahiz eta proteina ugari berez, inolako laguntzarik gabe eta katea osatzen duten aminoazidoen eta ingurunearen ezaugarrien arabera soilik tolesten diren, badira egoera natiboa bereganatzeko txaperoien laguntza ezinbestekoa duten proteinak ere. Txaperoien lana funtsezkoa izaten da hain beteta dagoen zelula barneko ingurunean proteinen artean gerta daitekeen agregazioa ekiditeko edota gehiegizko beroaren edo beste edonolako kondizio estresagarrien ondorioz proteinek jasan dezaketen toleste desegokia edo agregazioa ekiditen laguntzeko.
Ohikoak dituzten tenperatura-tarteetatik gora edota behera, zeluletako proteinak ezegonkortu egiten dira, eta destolestu edo desnaturalizatu egin daitezke (arrazoi horregatik beragatik igaroko da arrautzaren zuringoa, irakitean, gardena izatetik zuri opakua izatera). Solutu-kontzentrazio altuak, muturreko pH-ak, indar mekanikoak edota desnaturalizatzaile kimikoak egoteak gauza bera ekar lezake. Guztiz desnaturalizatutako proteina batek bere bigarren eta hirugarren mailako egiturak galduko ditu, eta zorizko kiribil gisa agertuko da. Kondizio jakin batzuetan, zenbait proteina birnaturalizatu egingo dira, eta jatorrizko tolesturara itzuliko dira; beste batzuetan, berriz, desnaturalizazioa itzulezina izango da.
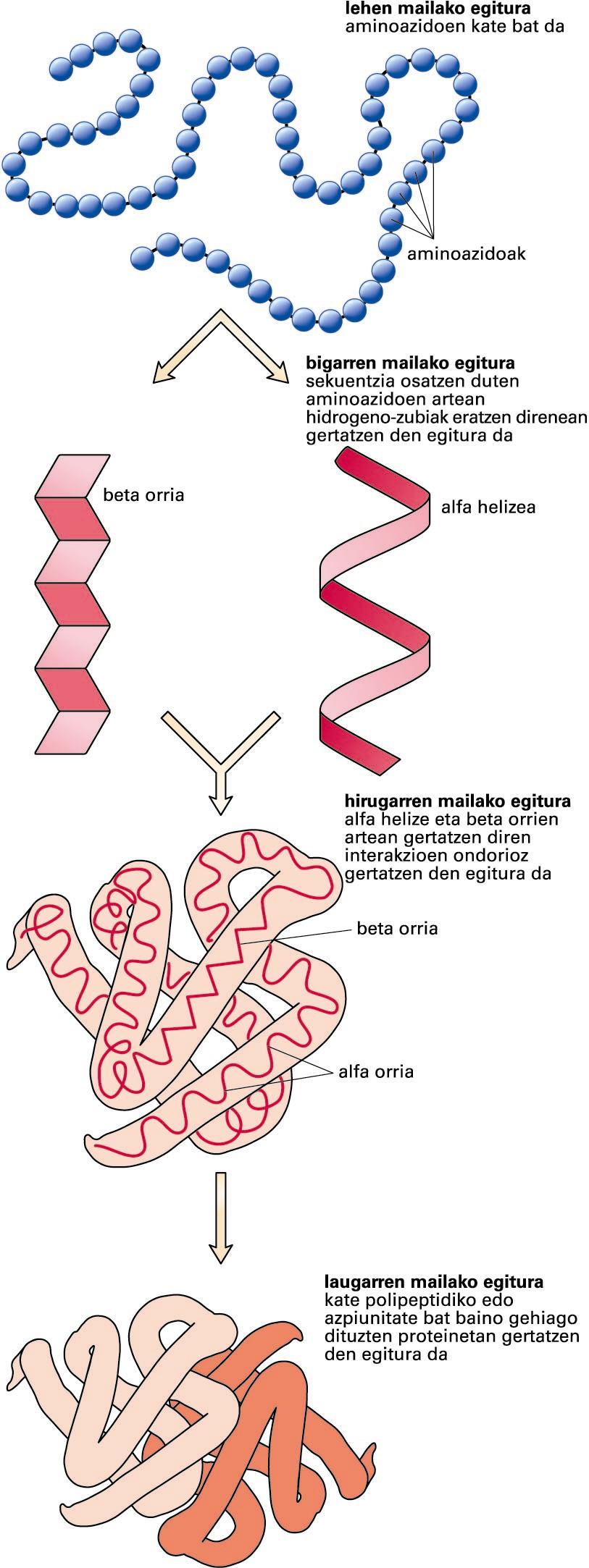
Proteinen egituraren lau mailak (iturria: National Human Genome Research Institute)
Proteinen egitura lau mailatan azaldu daiteke:
-
Lehen mailako egitura: aminoazido-sekuentzia. Lehen mailako egitura proteinaren sintesi-prozesuan eratzen diren aminoazidoen arteko lotura kobalenteek mantentzen dute. Lotura peptidiko horiek zurruntasuna ematen diote proteinari.
-
Bigarren mailako egitura: elkarren ondoko aminoazidoen artean eratzen diren hidrogeno-zubiz egonkortutako egiturak dira, eta aldizka errepikatzen dira. Adibiderik ohikoenak alfa helizea eta beta orria dira. Bigarren mailako egiturak proteinaren hainbat tokitan gertatzen direnez, molekula berean bigarren mailako egitura errepikatu eta desberdin asko aurkitu daitezke.
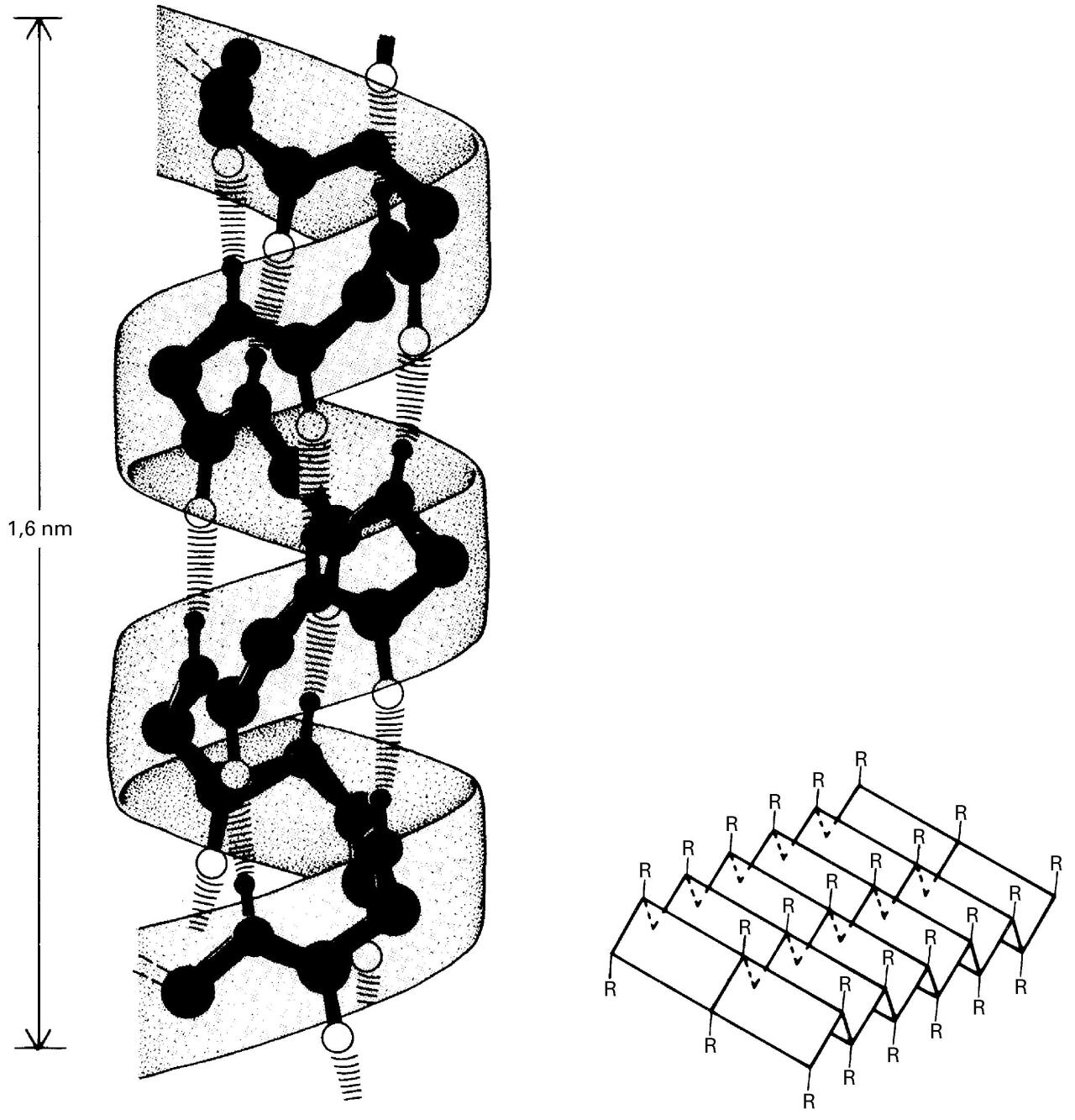
Bigarren mailako egituraren helizea (ezkerrean) eta orria (eskuinean)
-
Hirugarren mailako egitura: proteina-molekula baten egitura bere osotasunean; bigarren mailako egiturek euren artean espazioan hartzen duten forma. Hirugarren mailako egitura muin hidrofoboa eratzean egonkortuko da, baina egonkortze horretan zerikusia izango dute eratuko diren gatz-zubiek, hidrogeno-zubiek, disulfuro-zubiek edota itzulpen ondoko aldaketek.
-
Laugarren mailako egitura: proteina baten bi kate polipeptidiko edo azpiunitateren edo gehiagoren arteko interakzioaren ondorioz gertatzen den egitura. Egitura nagusiari proteina oligomeriko deritzo.
Baina proteinak ez dira molekula zurrunak. Aipatutako egitura-maila horietaz gain, antzekoak diren egitura batetik bestera pasa daitezke proteinak euren funtzioa betetzen ari diren bitartean. Berrantolaketa funtzional horri buruz ari garenean, hirugarren eta laugarren mailako egiturei konformazio deritze, eta konformazio desberdinen arteko trantsizioari, konformazio-aldaketa. Entzimetan, adibidez, substratua gune aktibora lotzeak konformazio-aldaketa dakar maiz. Disoluzioan proteina guztiek jasaten dute aldaketatxoren bat egituran, bibrazio termalaren edota ondoko molekulekin gertatutako talken eraginez, esaterako.
Proteinen hiru dimentsioko egituraren arabera, bi talde bereiz ditzakegu: proteina globularrak eta zuntz-proteinak. Proteina globularrek itxura esferikoa dute, gehienak disolbagarriak dira eta asko entzimak dira. Proteinaren talde apolarrak (isats hidrofoboak) molekularen muinerantz kokatzen dira, eta talde polarrak (buru hidrofiloak), ostera, kanporantz; hala, disolbatzailearekin dipolo-dipolo motako elkarrekintza gertatzen da. Horixe da hain zuzen ere molekularen disolbagarritasunaren arrazoia. Zuntz-proteinak egituretan azaltzen dira, eta harizpi luzeak eratzen dituzte. Normalean, egituretan azaltzen dira, edo biltegi-funtzioa duten proteina geldoak izan ohi dira. Disolbagaitzak dira gehienetan, eta agregatu trinko itxurakoak, molekularen kanpoalderantz begira kokatzen diren talde hidrofoboak direla eta. Ohikoak ez diren bigarren mailako egiturak izan ditzakete, kolagenoaren helize hirukoitza esaterako. Maiz, harizpien arteko crosslinkak dituzte, keratina-kateen arteko cys-cys disulfuro-zubiak adibidez. Zuntz-proteinek ehun konektiboa, tendoiak, hezur-muina edo giharretako muskuluak osatzen dituzte. Adibiderik argienak keratina, kolagenoa eta elastina dira.
Proteina globularrak zuntz-itxurakoak baino errazago desnaturaliza daitezke.
Aipatzekoak dira, talde gisa mintz-proteinak; zelularen edo organulu baten mintzari lotuta dauden edo harekin interakzioa duten proteinak dira. Bi motatako mintz-proteinak bereiz ditzakegu: mintz-proteina integralak eta periferikoak. Mintz-proteina integralak lipido-geruza bikoitzari lotuta daude beti. Proteina horiek mintzetik askatzeko detergenteak (SDS-a edo X-100 Tritoia) edo disolbatzaile apolarrak erabili behar dira. Integralen artean, mintza zeharkatzen duten proteinak ditugu batetik, eta integral monotopikoak, mintzari alde batetik soilik lotzen zaizkionak, bestetik. Proteina periferikoak kobalenteak ez diren elkarrekintza hidrofobo eta elektrostatiko bidez lotzen zaizkie lipido-geruza bikoitzari edo proteina integralei. Proteina horiek mintzetik askatzeko, pH altuko edo gatz-kontzentrazio handiko disoluzioak behar dira.
Mintz-proteinek itzulpenaren ondorengo aldaketak —gantz-azido edota GPI (glukosilfosfatidilinositol) aingurak— izan ditzakete.
Mintz-proteinak
Proteina baten hirugarren mailako egitura edo hark osatzen dituen konplexuen laugarren mailako egitura aurkitzea baliagarria izan daiteke proteina batek nola funtzionatzen duen argitzeko. Egituraren gaineko ikerketarako X izpi kristalografia eta NMR espektroskopia izeneko teknikak dira nagusi; atomo mailako bereizmena dute biek. Aurkitutako egiturak eskuragarri daude Protein Data Bank izeneko datu-basean. Hala ere, gaur egun, askoz gehiago dira gene-sekuentzia ezagunak proteina-egitura ezagunak baino. Horretaz gain, ezagutzen diren egituretatik gehientsuenak kristalizazio errazeko proteinei dagozkie. Proteina globularrak, esaterako, kristalizatzeko errazak dira, oro har; mintzproteinen kristalak lortzea, aldiz, zaila da, eta ondorioz, gutxienekoak dira Protein Data Banken. Proteinen egituraren iragarpen-metodoak baliagarriak izan daitezke esperimentalki zehaztea ezinezkoa den proteinen egiturak iragartzeko.