cell
- 1. Biol.
Izaki bizidunen unitate estruktural eta funtzionala; bizitzat jo ohi den unitate txikiena da.
- 2. Elektron.
Erradiazio-energia energia elektriko bihurtzen duten hainbat gailuri ematen zaien izena, hala nola eguzki-zelula edo zelula fotovoltaikoa.
- 3. Elektron./Fis./Kim./Teknol. Elektr.
- sin. voltaic cell, electrochemical cell, galvanic cell, electric cell
Erreakzio batean askaturiko energia kimikoa energia elektriko bihurtzen duen gailua.
- 4. Inform.
Kalkulu-orri batean edo datu-base bateko taula batean, zutabeen eta errenkaden arteko gurutzagunea, informazioak eta datuen arteko loturak jasotzeko erabiltzen dena.
- 5. Telekom.
Telefonia zelularrean sare zelularra osatzen duten geografia-unitate hexagonaletako bakoitza.
1. Biol.
-
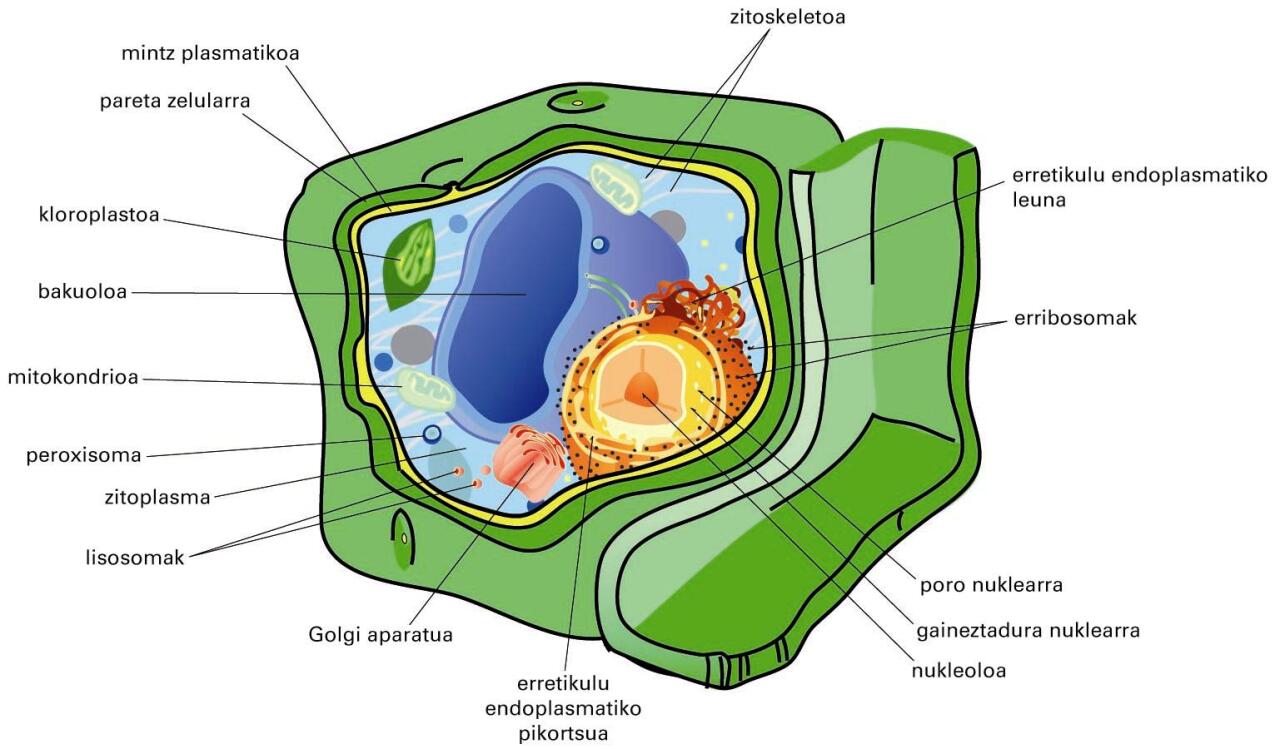
Landare-zelularen irudi eskematikoa. Ikus paretaren garrantzia eta zitoplasma urria (bakuolo erraldoia bertan baitago) -
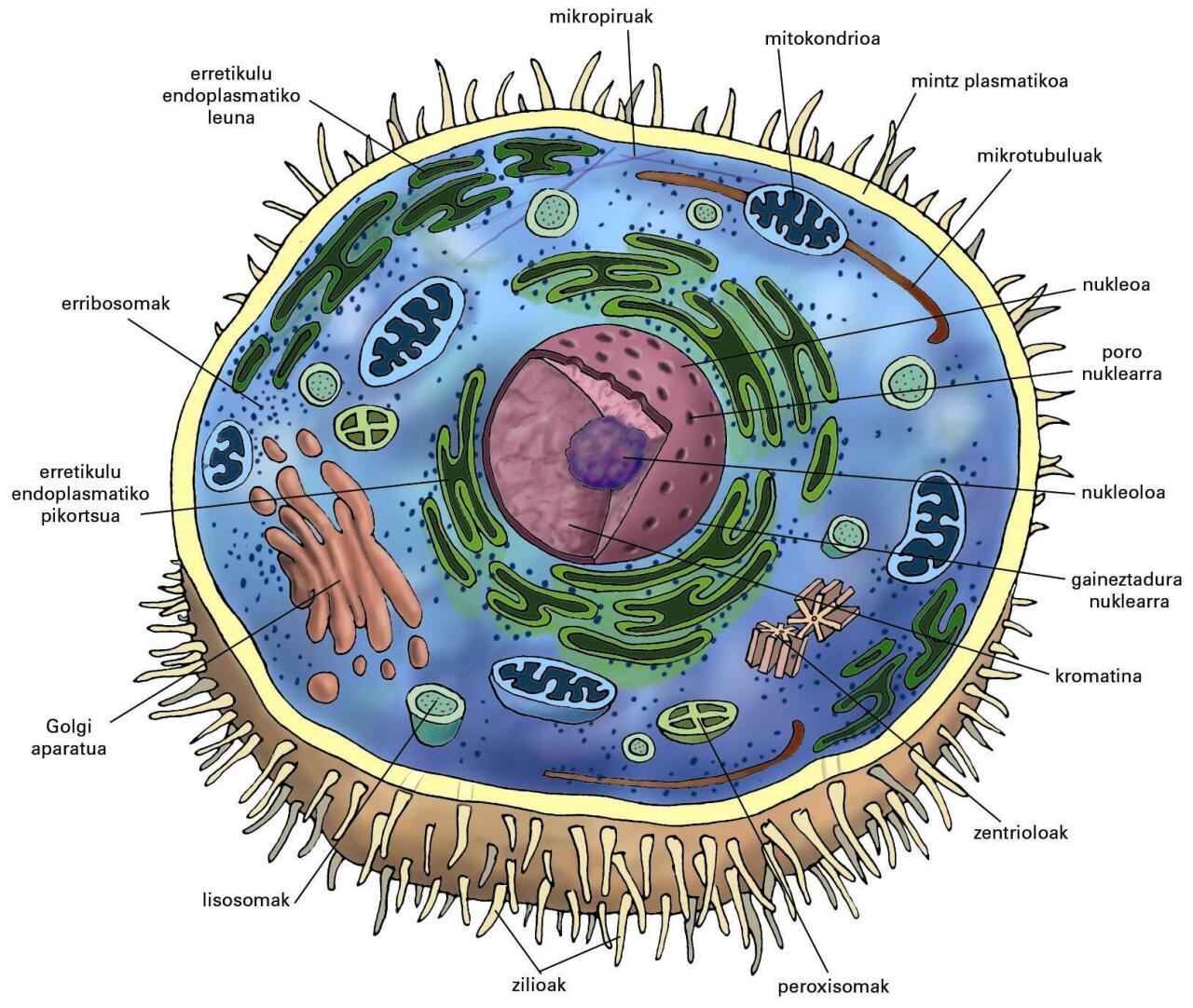
Animalia-zelularen irudi eskematikoa. Mintzari atxikirik, zilioak eta mikrobiloskak beha daitezke. Zelula hau ez dago polarizatuta (ez da epitelio-zelula) eta ezin dira eskualdeak (erpinaldea eta oinaldea ...
3. Elektron./Fis./Kim./Teknol. Elektr.
-
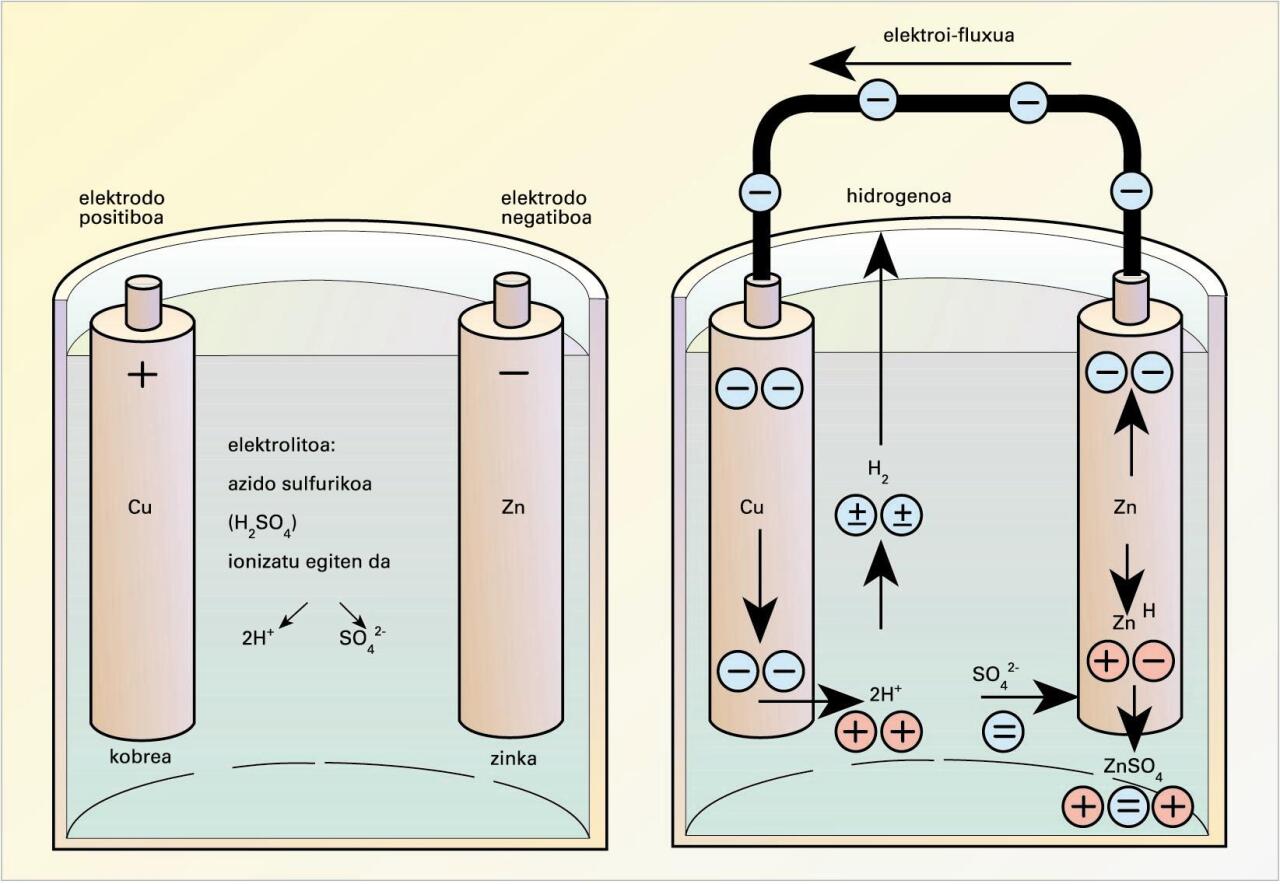
Daniellen pilaren eskema -
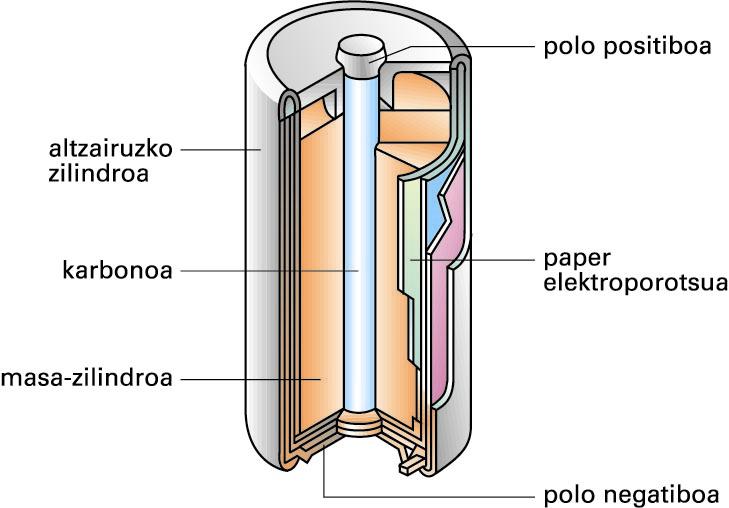
LeclanchÈren pila lehorra
- 1. Biol.
- Izaki bizidunen unitate estruktural eta funtzionala; bizitzat jo ohi den unitate txikiena da.
Zelula Edit
Egilea: Manuel Soto
Zelula (cellula, latinez) izaki bizidunen oinarrizko unitate morfologikoa da, eta, teoria zelularrekin bat eginez, bizitzat jo ohi den unitate funtzionala. Organismo unizelularren existentziak egiaztatzen du zelula dela unitate anatomiko sinpleena. Organismo unizelularrez gain, plurizelularrak daude, zelula anitzez osatuak. Egiturari erreparatuz, zelula guztiak oinarrizko bi patroitan antolatzen dira: prokariotoak eta eukariotoak. Behean azalduko dira xehetasun gehiagorekin, baina beren arteko desberdintasun nagusia da zelula eukariotikoek barne-mintzen sistema dutela. Zelula eukariotikoak bi motatakoak izan daitezke, animalia- eta landare-zelulak, alegia.
Zelula kontzeptuaren eboluzioa
Hastapenak
Zelula kontzeptua asko aldatu da mendeetan zehar, batez ere, berrikuntza teknologikoei esker. Mikroskopioa izan da lehenengo ezinbesteko tresna zelulak behatu ahal izateko. Janssen anaiek asmatu zuten 1595ean, baina garai hartan mikroskopioak jostailuak ziren, eta gortesauen aisialdia asetzeko gehien erabiltzen ziren laginak putzuetako ur ustela, odola eta beste isuri biologikoak ziren. Leuwenhoek izeneko mikroskopista ospetsuaren garai hartako irudiak baliagarriak ditugu: gaur egun museoetan dauden baina oraindik erabil daitezkeen mikroskopioak fabrikatzeaz aparte, zelulen marrazkiak (zelula hitza aipatu barik) argitaratu zituen xehetasun handiarekin (protozooak, espermatozoideak, bakterioak, etab). 1640. urtean, Stellutik euliaren anatomia mikroskopikoa argitaratu zuen. 1640 eta 1660 bitartean, Falopiok eta Glissonek proposatu zuten bizidunen oinarrizko unitatea zuntza zela.
Zelula hitza Robert Hooke ikerlari ingelesak 1665an argitaratu zuen Micrographia izeneko liburuan erabili zuen lehendabizi. Hookek gelaxka (cellula) hitza erabili zuen kortxoan eta beste zenbait landare-ehunetan mikroskopioa erabiliz ikusi zituen hutsuneak izendatzeko. Dena den, zelularen zati bat baino ez zuen ikusi, pareta zelularra, alegia. 1668an, Swammerdamek zelulak marraztu zituen lehen aldiz: odoleko globulu gorriak. Garai berean, Malpighik eta Grewek besikula izena eman zieten landare-zurtoinen ebakietan ikusten zituzten zelulei. Garai hartan, Grewek berak landu zuen ehun terminoa zentzu anatomikoan ornodunen zati elemental batzuk izendatzeko.
Teoria zelularraren planteamendua
1839an, bi ikerlari alemanek proposatu zuten teoria zelularra, Matthias Schleiden botanikariak eta Theodor Schwann zoologoak. Teoria horrek bi ideia orokor zeuzkan: batetik, bizidunen antolaketa, eta, bestetik, zelulen jatorria. Lehenengoa ez zen originala, garaiko giroan baitzegoen. Bigarrena, berriz, gakoa zen garrantzi handia eman zielako orduan ezagutzen ziren nukleo- eta nukleolo-motako egiturei. Zelan garatu zen teoria zelularra? Laburbilduz, 1838an, Schleiden teoria garatzen ari zen landareen zelulentzat baliagarria izan zitekeelakoan, eta haren ideiak Schwannekin konpartitu zituen. Schwannek animalien zelulen nukleoan egitura analogoak behatu zituen, eta biek hasi ziren zelula guztien batasunaren printzipioa eztabaidatzen. Schleiden eta Schwannen ideia horiek hasiera eman zioten teoria zelularraren planteamenduari, eta, 1839an, bizidun guztietarako baliagarria izan nahi zuen teoria zelularra proposatu, garatu eta defendatu zuten. Teoria zelularra honako bost postulatuotan oinarritzen da:
-
Zelula, bizidun guztien zati elemental edo unitate estrukturala da.
-
Zelulak hiru osagai nagusi ditu: mintza, gorputz zelularra eta nukleoa.
-
Izaki plurizelularren zelula bakoitzak bizitza bikoitza agertzen du: zelularena berarena eta izakiarengan txertaturiko elementua izateagatik dagokiona.
-
Teoria zelularraren oinarrizko arazoa zelulen jatorriari buruzkoa da.
-
Aurreko postulatuen gainean, de novo eraketaren hipotesia postulatzen da (masa organiko amorfo batean kondentsatu eta garatzen den nukleo batetik sortutakoa dela zelula).
Azkeneko puntuan proposatu zenak azalpen txiki bat merezi du. Schleidenentzat nukleoa zen berez sortzen zena eta hazi ostean bere baitan nukleoloa garatzen zuena. Azkenik, nukleolo horrek beste zelula berri bat sortzen zuen ustez. Schwanen ideia antzekoa zen, baina nukleoaren sorrera, endogenoa (Schleiden) izateaz gain, exogenoa izan zitekeen, hau da, nukleoa zelula amaren zitoblasteman (gai organiko amorfoa) sor zitekeen.
Teoria zelularraren ondorengoak XIX. mendera arte
Garai hartako kritika nagusia zitoblastemaren proposamena zen, berezko sortzapenarekin guztiz bat zetorrelako; ordurako, teoria hori nahiko kolokan zegoen. Horretaz gain, beste kritika garrantzikoa zatiketa zelularraren ingurukoa zen. Horren haritik, handik hemezortzi bat urtera (1858), Virchow ikerlari alemaniarrak zelula guztiak beste zelula batetik etorritakoak zirela (Omnis cellula e cellula) proposatu eta frogatu zuen. Hori dela medio, eta Louis Pasteurren garaiko ikerlariaren lanei esker, betiko errefusatu zen berezko sortzapenaren ideia. Hurrengo urteetan, zatiketa zelularreko mekanismoak deskribatu ziren: mitosiaren deskribapen zehatza (Walter Fleming, 1882), kromosomen deskribapena (Strassburger, 1884) eta abar.
Hala ere, bazeuden teoria zelularraren bestelako puntu ilunak ere; hala nola mintz zelularraren kontzeptua. Mintz zelularra ezin da mikroskopio optikoaren bitartez behatu; orduan, izen horrez izendatu zen egitura pareta zelularrari zegokion. Beste puntu ahula: teoriatik at gelditu ziren antolaketa sinpleagoa zuten organismoak, hala nola birusak, mikoplasmak, ricketssiak, klamidiak, bakterioak eta zianofizeoak, nukleo zelularrik gabeko izaki unizelularrak baitira. Gaur egun, izaki anukleatu horiek, birusak izan ezik, zelulatzat jotzen dira, eta prokarioto izena jarri zaie. Birusak autorreplikaziorako gai ez diren izaki zelularrak direnez eta beste zelula baten baitan ugaldu beharra dutenez, bizkarroitzat jotzen dira.
Gaur egun, ez da zalantzan jartzen neuronen izaera zelularra, baina XIX. mendearen bukaeran eztabaidagaia zen. Nerbio-sistema sare jarraitua da, eta garai hartako askorentzat (Köelliker ospe handiko mediku alemana barne) ez zen teoria zelularraren barruan sartzen; hau da, indibidualtasunaren ideiaren kontra zegoen. Santiago Ramón y Cajal histologoak, Golgi italiarrak garatu zuen zilarrezko inpregnazioaren teknika aplikatuz, neuronen osagaiak agerian jarri zituen (dendritak eta gorputz zelularra), eta bilbe konplexu hori zelula indibidualez osatuta zegoela frogatu zuen; bide batez, gainera, zelula-mota guztietarako teoria zelularra orokortu zen. Horren ostean, hainbat organulu zelular deskribatu ziren, besteak beste, Golgi aparatua, erretikulu endoplasmatikoa eta mitokondrioak (bai animalia-zeluletan eta bai landare-zeluletan ere).
Zelula kontzeptua XX. mendean
Lehenik eta behin, aipatu beharra dago XX. mendean berrikuntza handiak lortu zirela. Hasieran, genetistek eta biokimikariek zelulen barneko unitate txikiago bat bilatu nahi zuten, eta, aldi berean, zitologoek zelulen funtzionamendua deskribatzen jarraitu zuten. Kontzeptu horien garapenaren bitartez, aurrerapen teknikoak eskuratu ziren, hala nola kromatografiak, elektroforesiak, ultrazentrifugazioak, frakzionamendu zelularrak, in vitro analisiak, mikroskopia elektronikoa eta ikuspegi morfologiko berria (ultrastruktura), karakterizazio biokimiko zehatza eta abar. 1950eko hamarkadan, zenbait ikerlarik (haien artean, Claude, Palade, Porter eta De Duve) mikroskopia elektronikoa eta frakzioen azterketa biokimikoak bateratu zituzten, eta biologia zelular modernoa jaio zen. Garai hartan deskribatu eta karakterizatu ziren lisosomak, peroxisomak eta bestelako egitura zitosolikoak (mikropiruak, tonopiruak, mikrotubuluak) eta haien zeregin zelularra. Aldi berean, zilio, flagelo, gorputz basal, zentriolo eta ehoardatz akromatikoaren izaera mikrotubularra frogatu zen. Bukatzeko, esan beharra dago zelula eta ehunen in vitro kultiboen garapena ezinbestekoa izan zela biologia zelularraren arlo molekularra garatzeko.
Zelularen gaur egungo kontzeptua
Aurreko mendearen bukaeran eta honen hasieran gertatutako iraultza teknologikoaren barruan, hazkuntza zelularrak, azterketa genetikoak, teknika mikroskopiko berrien garapena, sistema informatikoak, prozedura immunologikoak eta beste zenbait teknika kokatzen dira. Horiek guztiek batera zelulen ikuspegi kimikoa eta morfologia uztartu dituzte. Dena den, zelularen biologia molekularraren gaur egungo nagusitasuna azaldu dugun historiaren ondorioa da. Teknika fisiko eta kimikoen bitartez, zelulen baitan gertatu ohi diren zenbait fenomeno biologiko eta maila molekularrean gertatzen diren asoziazioak ulergarri bihurtu dira. Hala ere, mendeetan zehar garatutako kontzeptuak, tresneriak eta hurbilketa teknologikoak zelula unitate elementaltzat jotzera eramaten gaitu.
Zelularen deskribapena
Arestian aipatu dugun bezala, zelula guztiak oinarrizko bi patroitan antolatzen dira: zelula prokariotikoak eta zelula eukariotikoak.
Zelula prokariotikoa
Antolakuntza prokariotikoaren morfologia oso sinplea da, baina aniztasun metaboliko handia adierazten du. Prokarioto gehienek pareta zelularra dute mintz plasmatikoaren kanpoaldean. Eubakterioa D-aminoazidoz osatutako mureina izeneko proteinaz egina dago. Izaera hori oso bitxia da, zeren munduan zehar aurkitu ditzakegun proteina guztiak L-aminoazidoz osatuta baitaude. Arkeobakterioek D-amoinoazidoez osatuta dagoen pseudomureina izeneko proteina dute. Paretaren funtzioa estrukturala da, eta presio osmotikoaren aldaketei aurre egiteko eta gaiak iragazteko balio du. Paretaren azpialdean, mintz plasmatikoa (edo plasmalema) dago. Mintz horren desberdintzapena ez da oso handia, eta ez du tolesturarik. Salbuespena fotosintesia egiten duten zianobakterioak dira. Prokarioto guztietan, tolestura xume bat dago mintzean txertatuta, mesosoma, hain zuzen ere. Hor, kromosoma eta erreplikazio-tresneria ainguratuta daude. Prokariotoen barrunbeari zitosol deritzo; hor, tresneria entzimatiko guztia dago, eta erribosomak ere badaude. Erribosoma horiek eukariotoenak baino txikiagoak dira (55 proteina eta 3 rRNA-molekulaz osatuak) eta askoz ugariagoak (Escherichia coli heste-bakterioaren kasuan, 15.000 aletik gora daude). Zitosolean (eta mintzari atxikia), DNA-molekula luze (1 mm) eta zirkular bat dute. DNAren etengabeko erreplikaziorako mintzean txertatuta, erreplisomak daude, eta transkripzioa eta erreplikazioa gertatzen dira aldi berean. Genoma horren garrantzia bere plastikotasunean datza. Hau da, zelulek geneak truka ditzakete zenbait prozesuren bitartez. Prokariotoetan ez dago zitoskeletorik, eta zatiketa zelularra mintz plasmatikoaren ildo batean gertatzen da. Zatiketa orekatua gertatzen da, eta, hori dela eta, zelula haurrideek kromosoma bana daukate.
Zelula eukariotikoa
Animalia, landare, onddo eta protisto plurizelular guztiak dira eukariotoak. Beraz, aniztasun handiko antolakuntza zelularra daukagu, eta zaila da eskema komunaz hitz egitea. Dena den, zenbait orokortasun aipa daitezke. Eskuarki, zelula eukariotiko guztiek egitura bera dute, eta lau elementuz osatuta daude: kanpoaldea, konpartimentuen muga, barnealdea eta genoma.
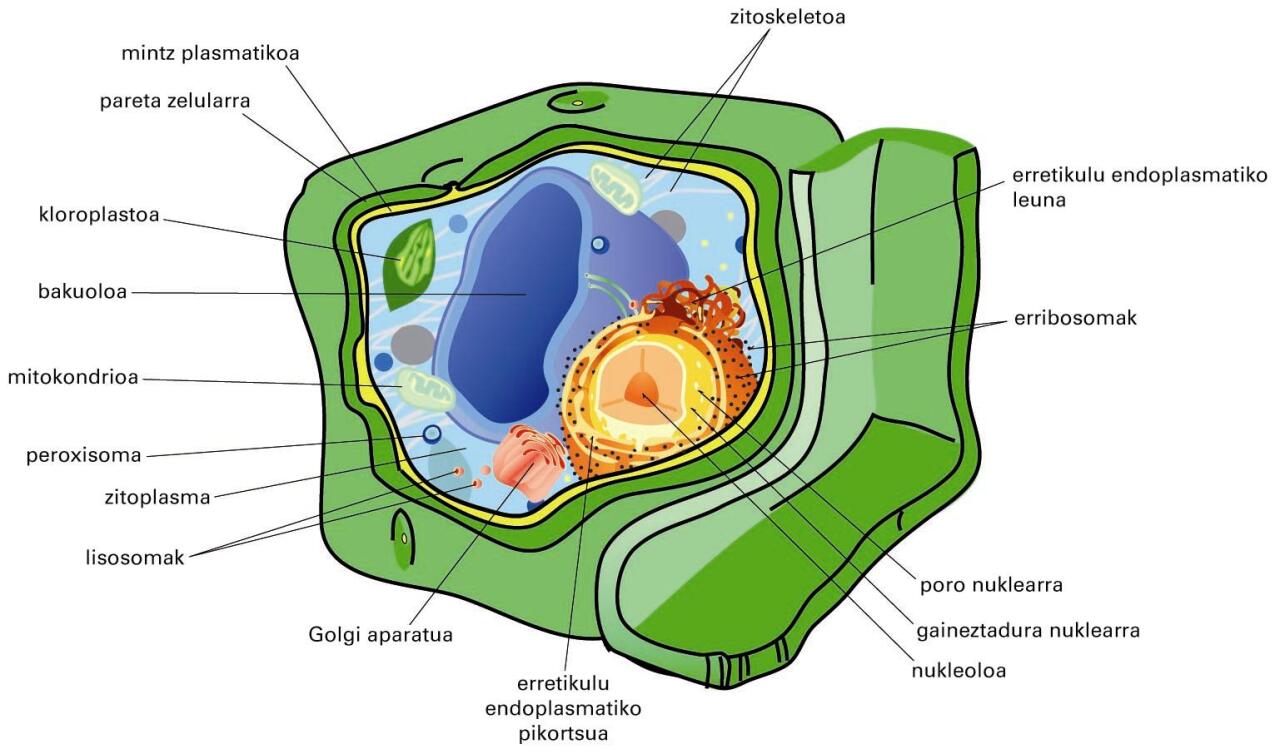
Landare-zelularen irudi eskematikoa. Ikus paretaren garrantzia eta zitoplasma urria (bakuolo erraldoia bertan baitago)
Kanpoaldea edo zelulaz kanpoko matrizea
Ingurune zelularrari dagokio, eta seinale-molekulez zein uraren orekaz eta zelularen eusteaz arduratzen diren zelulek berek jariaturiko zuntz-proteinez eta makropolisakarido erraldoiez osaturik dago. Matrizea ez da substantzia inertea; alderantziz, funtzio aktibo asko betetzen ditu. Adibidez, zelulen zitoskeletoarekin erlazio estuan egoteagatik zelulen jokaera eraentzen du. Zelulara iritsi behar duten substantzia eta konposatu guztiek zeharkatu behar dute zelulaz kanpoko matrizea, eta haien iragaitea eta iragaite-abiadura kontrolatzen ditu. Beraz, zelulen garapena, funtzio metabolikoak, desberdintzapena, proliferazioa, forma eta migrazioa matrizearen menpekoak dira. Behean aipatuko diren zitomintz-sistemen eta organulu zitoplasmatikoen mintzak morfologikoki mintz plasmatikoaren antzekoak dira. Hortik dator mintz unitarioaren kontzeptua. Funtzionalki desberdin samarrak izan daitezke, organulu edo zelula-motei dagokien funtzio espezifikoa bete ahal izateko. Adibidez, mitokondrioaren barne-mintzean proteinak nagusi dira, energiaren transdukzioarekin (ekoizpena) zerikusia daukaten prozesuak egiteko, eta neuronen mintzaren konposizioa lipidoz aberatsa da, iragazkortasuna mugatzeko eta nerbio-bulkada modu eraginkorrean transmititu ahal izateko.
Konpartimentuen muga (barrera biologikoa) edo mintz plasmatikoa
Lipidoz (iragazkortasunerako), proteinaz (garraiorako) eta karbohidratoz (informazio-trukerako) osatua dago. Kanpo-ingurunearekin kontaktuan dago, eta, iragazkortasun selektiboari esker, zelulen arteko zein zelula eta kanpo-ingurunearen arteko trukeaz arduratzen da. Zelularen kanpoalde eta barnealdearen arteko ioien gradienteak mintzari esker sortu eta mantentzen dira. Beraz, mintzen funtzio garrantzitsuenetakoa kanpotik datozen seinaleak ezagutzea da, eta seinale horiei erantzun egokia ematea. Landare, alga eta onddoen zelulen kanpoaldean, mintz plasmatikoaren gainetik, egitura zurruna dute, hau da, pareta zelularra (ikus aurreko irudia). Egitura horren osagaiak polisakaridoak, monosakaridoak eta gatz mineralak dira, baina osagai nagusia zelulosa da (glukosa-polimero bat). Landare-zelulen paretaren funtzioa ez da soilik estrukturala, landarearen formaren euskarria da, eta landarearen fluidoak garraiatzen eta babesten ere parte hartzen du.
Barnealdea, erreakziogunea edo zitoplasma (proteinak, entzimak eta abar)
Mintzez mugaturiko egiturez beteta dago: zitomintz-sistema (erretikulu endoplasmatiko leuna eta pikortsua, Golgi aparatua, jariapen-pikorrak, endosomak, lisosomak, gaineztadura nuklearra eta metatze- eta garraio-besikulak. Egitura horiek guztiak sistema bakar batean integratuta daude, eta lotura zuzenak edo zeharkakoak dituzte mintz plasmatikoarekin. Sistema konplexu eta tolesturadun honen presentziak zelularen tamainaren emendioa dakar, bakterioekin konparatuz gero, zeren eukariotoen azalera/bolumena erlazioa 1/20 ingurukoa baita. Zitomintz-sistemen barruan, erretikulu endoplasmatiko pikortsuak eta leunak daude. Lehenengoen funtzioa proteinen sintesia da, eta bigarrenena, lipidoena. Hor sintetizaturiko proteinak eta lipidoak mintzez inguratutako garraio-besikuletan barneratzen dira, eta Golgi aparatura bidaltzen dira helduak/aktiboak izateko beharrezkoak diren zenbait prozesu jasateko (glikosilazioak, sulfatazioak eta prozesamendu proteolitikoak, alegia). Golgi aparatutik lisosomak ere ekoizten dira. Mintz bakarra dute, eta beren barrualdean hidrolasa entzima azidoak dituzte, aktibo izateko pH azidoa behar dutenak. Lisosomen funtzioa beren barrualdera iristen diren substantziak digeritzea da.
Zelularen forma mantentzeko eta zitomintz-sistemaren osagaiak antolatzeko, piruz eta tubuluz osaturiko eskeletoa dago zelularen barnealdean: zitoskeletoa. Egitura hori unibertsalak diren aktinazko piruez eta mikrotubuluez osatuta dago. Biek motor biologiko gisa jardun dezakete, ATPasak diren proteinekiko elkarrekintzen bidez, besikulak eta pikorrak mugiaraziz. Unibertsalak ez diren beste osagaiak ere badaude: piru ertainak, alegia. Zitomintz-sistema ez den eremuari zitosol deritzo, eta proteinak, koentzimak eta metabolitoak daude hor.
Eukariotoek, zitomintz-sistemaz gain, mintzak mugaturiko organuluak dituzte: mitokondrioak, peroxisomak eta plastoak —azken horiek landare-zeluletan baino ez daude)—. Lehenengo biak mintz bikoitzez inguraturiko organuluak dira, eta barne-mintzetan energia (ATP) ekoizteko tresneria guztia dute (fosforilazio-sistemak). Organulu horien baitan, prokariotoen matrizearen antzeko eta izen bereko eremua dago. Eremu horretan, organuluek genoma nuklearrarengandik autonomoa den genoma propioa dute. Genoma hori DNA-kate zirkular batez osaturik dago, eta prokariotoek erreplikapen eta itzulpenerako duten tresneriaren antzekoa dute. Ikerle askok berezitasun horiek iraganeko prokariotoekin lotu dituzte, eta jatorri prokariotikoa ondorioztatu dute. Peroxisomak eta erlazionaturiko beste organulu energetikoak (glioxisomak, glikosomak, etabar) mintz bakuneko organuluak dira, eta, mitokondrioek eta plastidoek ez bezala, ez dute genoma autonomorik.
Genoma
Eukariotoetan benetako nukleoa agertzen da. Kromosoma prokariotikoarekin konparatuz gero, desberdintasun batzuk aurkitu ohi dira:
-
Eukariotoetan, DNA gaineztadura nuklearreko mintzari atxikia dago.
-
Prokariotoetan, informazio genetikoa DNA-harizpi bikoitz zirkular batean dago; eukariotoetan, berriz, gene zatikatuak dituen DNA-elementu lineal anitzetan.
-
Eukariotoetan, DNArekin batera, proteina paketatzaileak (histonak) daude, eta nukleosoma izeneko unitate morfofuntzionala osatzen dute. Beste antolakuntza-maila batean, nukleosomak beste elementu osagarriekin elkartzen dira kromatina eraikitzeko.
-
Eukariotoetan, transkripzioaren eta itzulpenaren arteko banaketa fisikoa dago, hau da, transkripzioa gune nuklearrean gertatzen da, eta itzulpena, zitosolean. Horri esker, RNA-molekulak zitosolera abiatzen dira, eta, nukleoloan (nukleoaren barruan) sintetizatu eta zitosolera migratu diren erribosomekin elkartzean, proteinen ekoizpena burutzen da. Proteinen sintesia zitosolean (erribosoma askeetan) edo erretikulu pikortsuan (mintzean txertaturik dauden erribosometan) gerta daiteke.
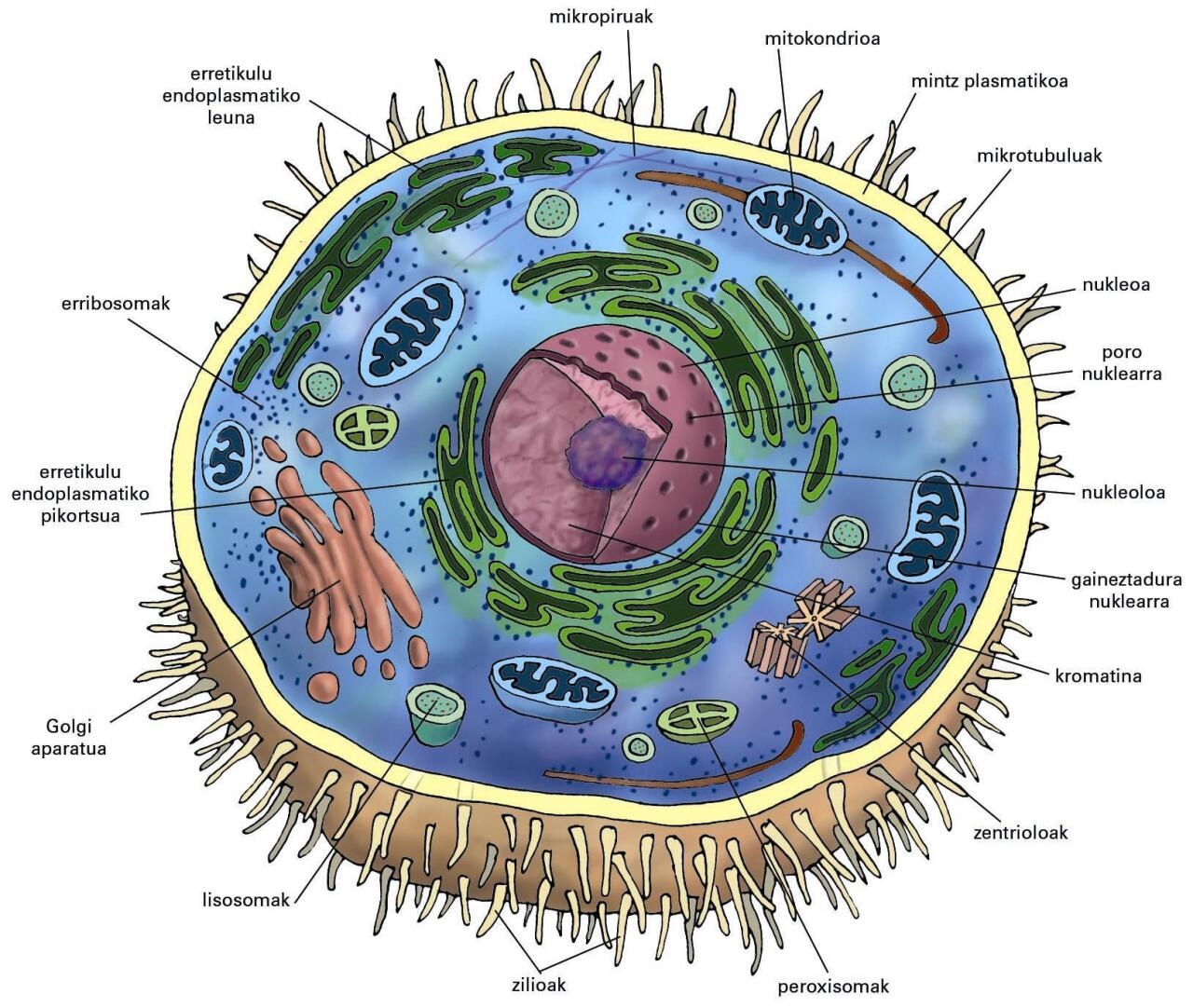
Animalia-zelularen irudi eskematikoa. Mintzari atxikirik, zilioak eta mikrobiloskak beha daitezke. Zelula hau ez dago polarizatuta (ez da epitelio-zelula) eta ezin dira eskualdeak (erpinaldea eta oinaldea) bereizi
- 3. Elektron./Fis./Kim./Teknol. Elektr.
- Erreakzio batean askaturiko energia kimikoa energia elektriko bihurtzen duen gailua.
Pila elektrokimikoa (edo pila) bere baitan gertatzen den erreakzio kimiko bati esker elektrizitatea lortzen duen muntaia esperimentala da. Sarri pilak eta bateriak sinonimotzat jo arren, bien artean egon badago diferentzia txiki bat.
Erreakzio kimikoak eta, horietan barne, oxidazio-erredukzio erreakzioak, erredox erreakzioak, itzulgarriak edo itzulezinak izan daitezke. Erreaktiboak soilik produktu bihur daitezkeenean, erreakzioa itzulezina da (produktuak ezin baitira erreaktiboetara itzuli); eta alderantziz, erreaktiboak produktu bihur daitezkeen bezala produktuak erreaktibo bihurtzen badira, erreakzioa itzulgarria da.
Horren kariaz, pila elektrokimikoetan gertatu ohi diren erredox prozesua eta muntaia esperimentala itzulezinak direnez, pilak ezin dira berriz kargatu; agortutakoan zaramara botatzen badira, kutsadura kimiko arriskutsua sortuko dute; pilak metatzeko gordailu berezietara eraman behar dira. Ostera, baterietan gertatzen diren erredox erreakzioak itzulgarriak direnez, neurri handi batean behin eta berriro erabilgarriak izan daitezke, berriz kargatzekoak baitira. Horretan datza, hain zuzen, aipaturiko diferentzia.
Pila batean gauzatzen den erredox erreakzioa bi erdielementutan banatu behar da muntaia esperimentala egiteko eta, horren bidez, korronte elektriko zuzena lortu ahal izateko, irudi honetan ikus daitekeenez:
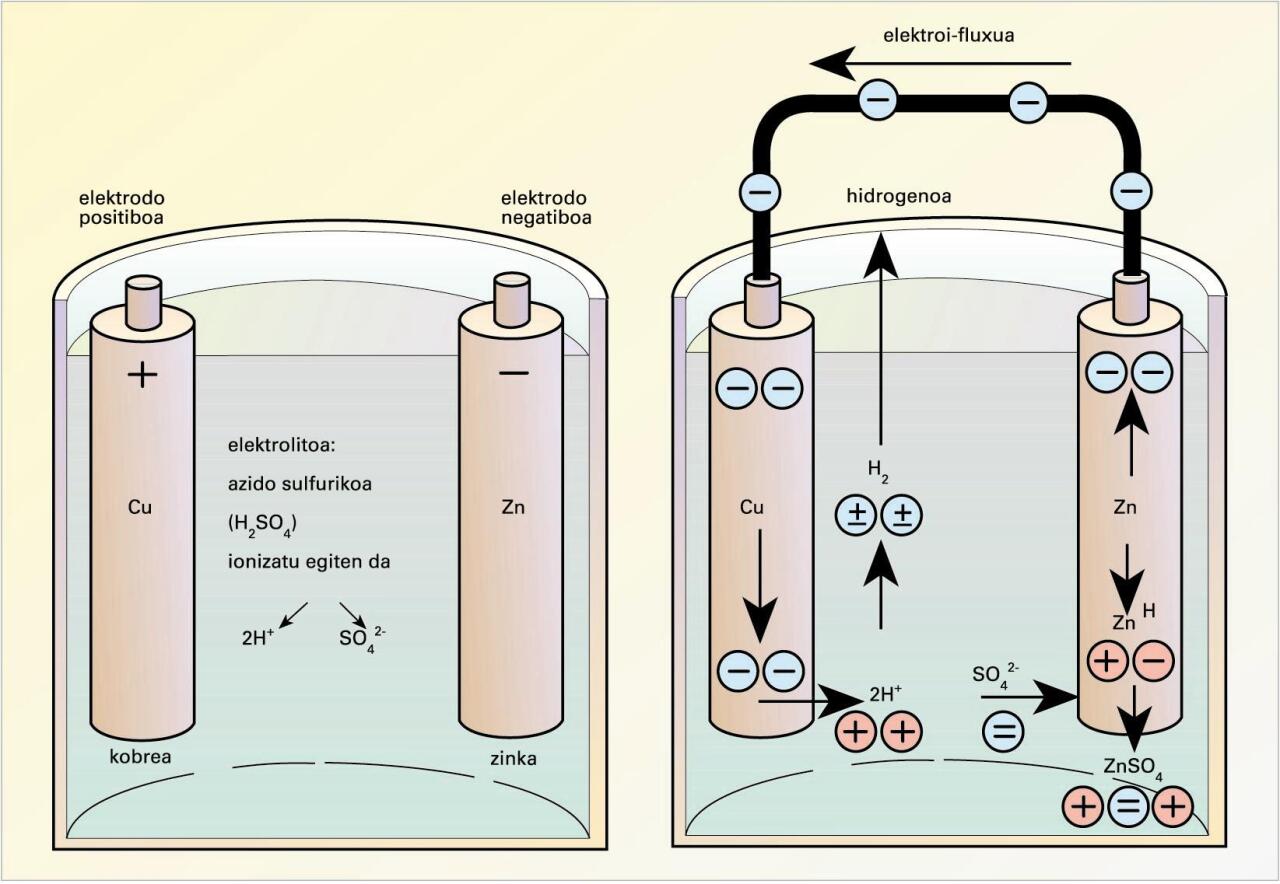
Daniellen pilaren eskema (iturria: EVE-EEEren Energiaren Hiztegi Entziklopedikoa)
Katodoan, erredukzioa gertatuko da, esate baterako, Cu2+(aq) + 2e− → Cu0(s), eta anodoan, oxidazioa, esate baterako, Zn0(s) – 2e−→ Zn2+(aq); erreakzioa denetara, Zn(s) + Cu2+(aq) Zn2+(aq) + Cu0(s).
Muntaia esperimentalean, metalak, era banatuan, beren ioien disoluzio ionikoetan murgilduta daude, eta metal biak eroale metaliko baten bidez lotuta. Halaber, disoluzio elektrokimikoek gatz-zubi baten bidez elkarri lotuta egon behar dute, disoluzioetan gerta daitezkeen kargen metaketak ekiditeko eta elektroiek kanpoko zirkuitu elektrikotik bidaiatu ahal izateko.
Esperimentalki neurtutako pilen tentsioak erreakzionatzaileen izaerarekiko, beren kontzentrazioarekiko eta tenperaturarekiko menpekotasuna dauka, Nernstek (1920) proposaturiko ekuazioan ikus daitekeenez:
Pila elektrokimiko “busti” horien desabantaila nagusia hau da: likidoak jariakorrak direla eta alde batetik bestera eramateko ez direla batere erosoak. Horregatik, garapen teknologikoa dela medio, antzeko pila lehorrak agertu dira merkatuan; Leclanchéren pila (1868) lehenetarikoa izan zen:
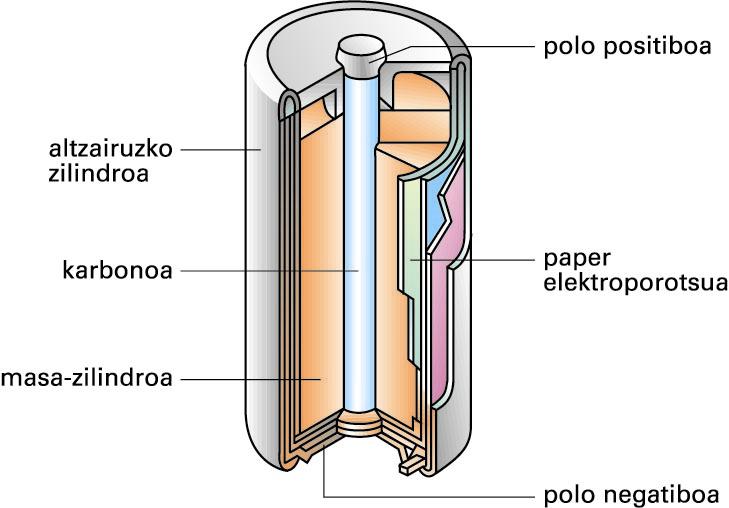
Leclanchéren pila lehorra (iturria: EVE-EEEren Energiaren Hiztegi Entziklopedikoa)
Horren ostean, pila alkalinoak, merkuriozkoak deiturikoak, litiozkoak eta abar merkaturatu dira, eta gaur egun horiek dira gure etxeko tresneriaren jaun eta jabe.
Erregai-pila deritzona etengabe kargatzen ari den pila bat da, erreaktiboak (erregaiak) etengabe hornitzen ari baitira erredox prozesuan, esate baterako, 2 H2(g) + O2(g) H2O(l). Eo = 1,23 V.
Pila horien abantaila nagusia zera da: hidrogenoaren errekuntza dela eta, etekina ia % 100 izan daitekeela; ostera, makina termiko batean, Carnotek etekina mugatzen du, zeina gutxi gorabehera % 33 den. Askoz energia erabilgarri gehiago lortzen da hidrogenoaren errekuntza prozesu elektrokimikoetan gertatzen bada, sugarretan gertatzen bada baino (erregai-pila).